![]()
Chapter 2:
Tick-borne flavivirus complex – phylogeny and biogeography
Gerhard Dobler
Key points
- Tick-borne flaviviruses are grouped in the genus Orthoflavivirus of the family Flaviviridae.
- According to the new taxonomic rules all orthoflaviviruses are named in a binominal way.
- 12 virus species of the genus Orthoflavivirus belong to the complex of tick-borne orthoflaviviruses, of which 6 are proven human pathogens.
- Tick-borne encephalitis virus (TBEV) is the medically most important tick-borne orthoflavivirus.
- Despite considerable progress in the understanding of tick-borne orthoflaviviruses and tick-borne encephalitis virus in detail, there are big gaps in the understanding of the evolution and biogeographic distribution of tick-borne orthoflaviviruses.
- The biogeographic distribution and spread of TBEV are still unsolved and with an ever increasing amount of data it becomes even more unclear.
Introduction
TBEVs belong to the family Flaviviridae. They are grouped in the newly established genus Orthoflavivirus and according to the modern taxonomy are now named in a binominal form. However, this modern taxonomic classification does not go into more detail and does not reflect epidemiological and pathogenetic aspects. Using TBEV as an example, it´s more detailed phylogeny and bio-geography are summarized here with the goal to better understand the natural history of these viruses. This may allow development of concepts for surveillance, viral spreading and emergence of infectious diseases caused by flaviviruses in general.
The tick-borne flavivirus complex
The family “Flaviviridae” is a group of viruses which have as common characteristics a typical spherical shape, a lipid envelope and a non-segmented single-strand RNA of positive polarity. In contrast to other virus families with a similar genome, the structural proteins are located at the 5’ end and the non-structural proteins are located at the 3’ end. Flaviviridaecontain a single open reading frame. The translational initiation is cap-dependent in the genus Orthoflavivirus while in the other genera it depends on internal ribosomal entry site elements.1 According to a recent re-classification and re-naming the family of Flaviviridaeis divided into 4 genera:
- Orthoflavivirus
- Pestivirus
- Pegivirus
- Hepacivirus
A total number of 89 virus species have been actually grouped into the family Flaviviridae.2 The medically most important genus of the family is the genus Orthoflavivirus. In the new binominal taxonomy of viruses the genus Orthoflavivirus contains a total of 53 viruses.2 The official taxonomy and classification do not differentiate into lower levels than species. However, the members of the actual genus Orthoflavivirus, former Flavivirus, are divided according to their transmission and phylogeny into three complexes.3
- Mosquito-borne flaviviruses,
- Tick-borne flaviviruses,
- non-vector borne flaviviruses.
The mosquito-borne complex of flaviviruses includes a number of important viruses of human pathogenicity, among them the type species of the family, according to the new taxonomy Orthoflavivirus flavi (former: yellow fever virus), the dengue viruses Orthoflavivirus dengue (no longer discriminating anymore between the four dengue virus serotypes and including them in one species), Orthoflavivirus japonicum (Japanese encephalitis virus), Orthoflavivirus nilense (West Nile virus including Kunjin virus), and Orthoflavivirus zikaense, (Zika virus). One big disadvantage of this new nomenclature system is that it is based only on genetic characteristics whereas epidemiological or transmission-based characteristics are no longer included.
The tick-borne Orthoflavivirus complex includes now a total of 13 flaviviruses (see Table 1). According to their epidemiological patterns and natural hosts two sub-complexes are distinguished (Table 1):
- Mammalian tick-borne flavivirus complex
- Seabird tick-borne flavivirus complex
Table 1: Current members of the tick-borne complex of the genus Orthoflavivirus
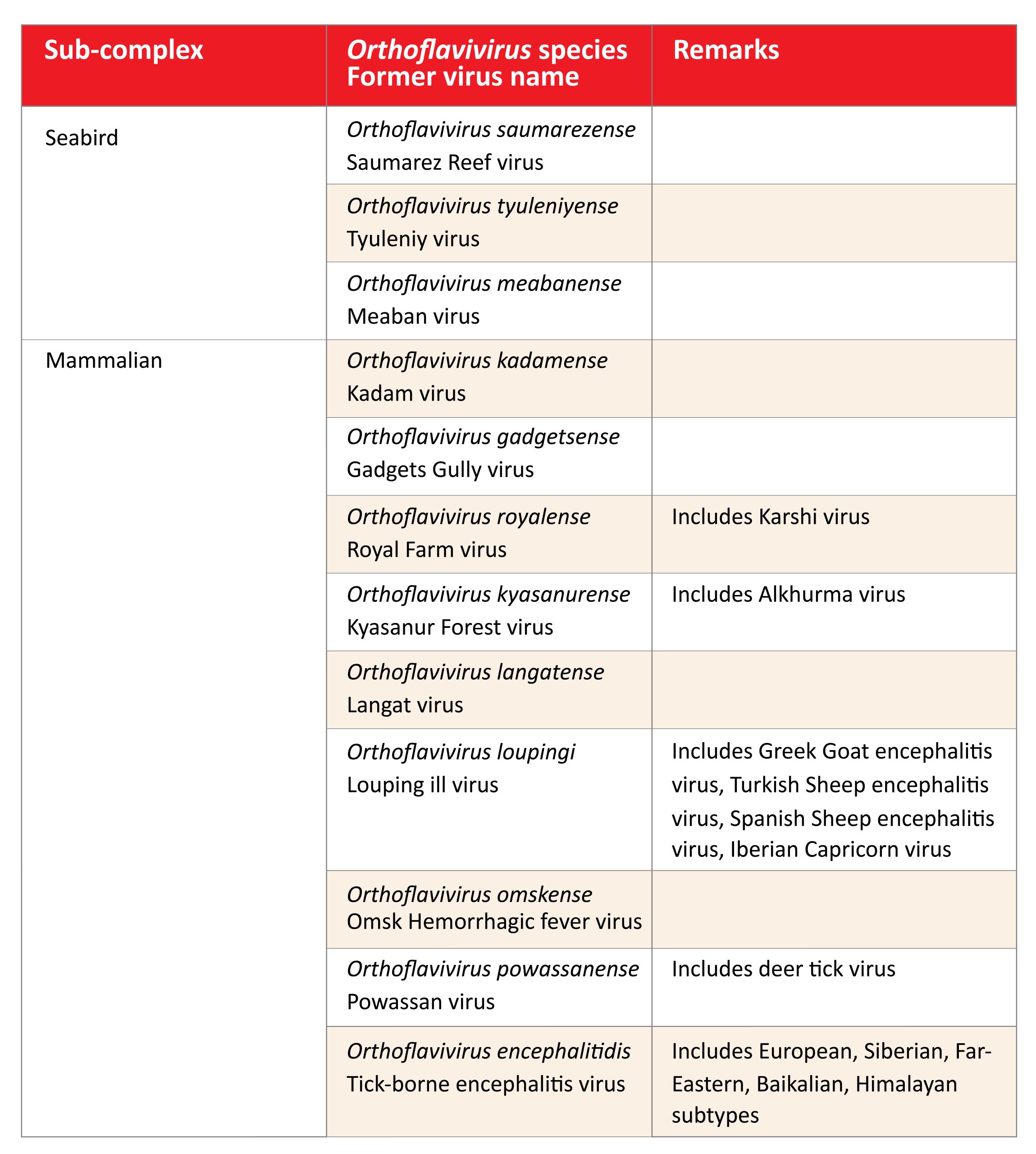
Click the image above to enlarge
Origin and spread of the Flavivirus tick-borne complex
According to the current concepts, tick-borne flaviviruses originated in Africa and are descendants of a pre-flavivirus which seems to be most closely related to Tamana Bat virus, a non-vector-borne flavivirus. Genetic analyses estimate that the orthoflaviviruses originated around 64,000 -160,000 years ago.4 The analyses also show that the mosquito-borne flaviviruses diverged from the tick-borne flaviviruses around 20,000 years ago,5 according to other analyses about 40,000 years ago4 and around 30,000 years.6 Kadam virus seems to be the first descendant of these very early tick-borne flaviviruses. It seems that already in this very early phase of evolution of tick-borne flaviviruses the seabird sub-complex of flaviviruses separated from the rest of the viruses, which finally developed into the group of mammalian sub-complex flaviviruses. The ways in which the three seabird-related flavivirus species spread on the shores of Europe, Asia and southern Pacific remains unsolved, but it can be speculated that migrating birds spread the viruses or virus-infected ticks to the different parts of the world (Fig 1).
It is assumed that the principle of tick-borne transmission of flaviviruses evolved in Africa, several tens of thousands of years ago. From there the tick-borne flaviviruses migrated in eastern direction to the Middle East where Alkhurma virus and Kyasanur Forest virus separated and evolved separately on the Arabian Peninsula and the Indian subcontinent respectively. Furthermore, Langat virus separated and migrated to the Malaysian Peninsula. The original virus dispersed to the northeast and finally reached the Siberian region,7 where it seems to have separated into the different subtypes of tick-borne encephalitis virus. In Siberia the Siberian subtype, the Baikalian subtype of tick-borne encephalitis virus and Omsk Hemorrhagic Fever virus and potentially even more (yet unknown) subtypes developed; further east the Far Eastern subtype of tick-borne encephalitis virus and the Himalayan subtype evolved. Powassan virus evolved and left the Asian continent to enter the North American continent. It is yet unclear whether the virus used the Beringian Landbridge (13,000-11,000 years ago) or if it was transported by migrating birds to this new continent.4 The exact mechanisms are unclear as is the relationship between the Russian and the American strains of Powassan virus. However, a recent study showed that the so-called lineage II of Powassan virus, Deer Tick virus, emerged between the 1940s and the 1970s in Northeastern U.S.A., possibly due to changes and adaptation of Powassan virus lineage I to new vector or host populations.8
Figure 1: Putative spread of viruses of the mammalian tick-borne flaviviruses according to different sources3-6
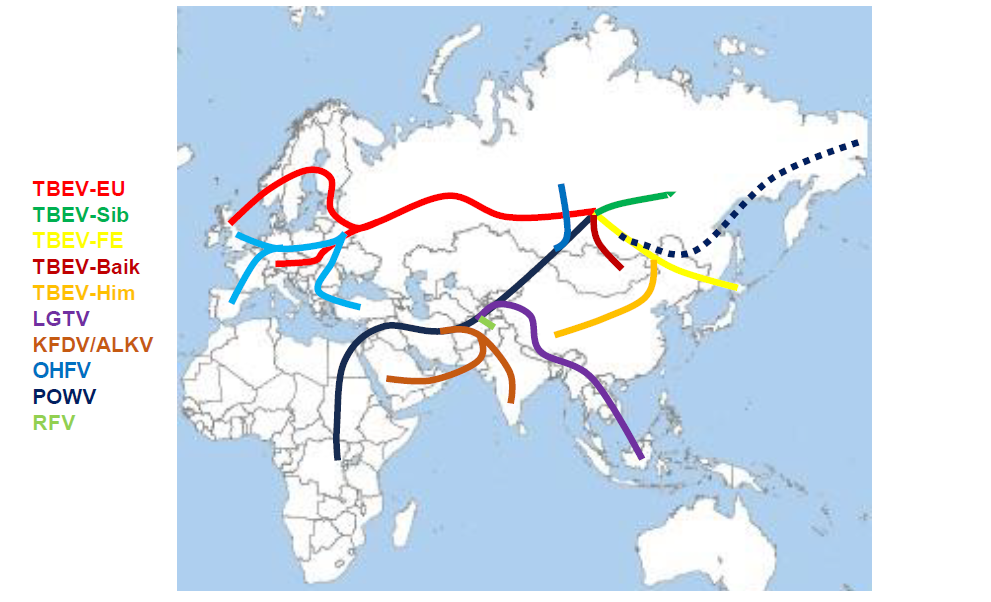
Click the image above to enlarge
The European subtype of tick-borne encephalitis split from the original virus probably somewhere in Siberia and migrated west to develop in the European subtype. There was another split to the south creating Greek Goat encephalitis virus and Turkish Sheep encephalitis virus and to the west to develop Louping ill virus on the British islands, Spanish Sheep encephalitis virus and the Iberian Capricorn virus, all subtypes of Louping ill virus. However, the exact way of distribution and spread remains to be better elucidated. The actual distribution of the members of the tick-borne complex of orthoflaviviruses is shown in figure 2.
Figure 2: Actual geographic distribution of the members of the mammalian tick-borne orthoflavivirus complex
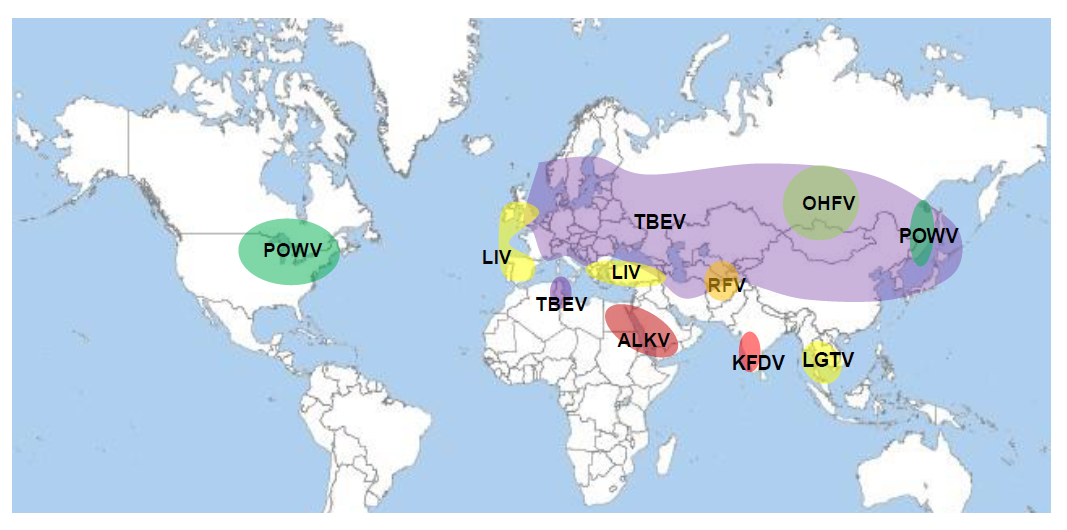
Click the image above to enlarge
Most members of the mammalian tick-borne flavivirus serocomplex have a human and/or a veterinarian pathogenic importance (Tab 2). In contrast, so far, no human or veterinarian disease could be associated with the members of the seabird tick-borne flavivirus complex. One can speculate that the specific viruses only have the potential to infect mammals and therefore humans when adapted to a mammalian natural host. It seems that the seabird tick-borne viruses are genetically not able to infect mammalian hosts.
Table 2: Members of the tick-borne Orthoflavivirus complex and main vector and pathogenic potential
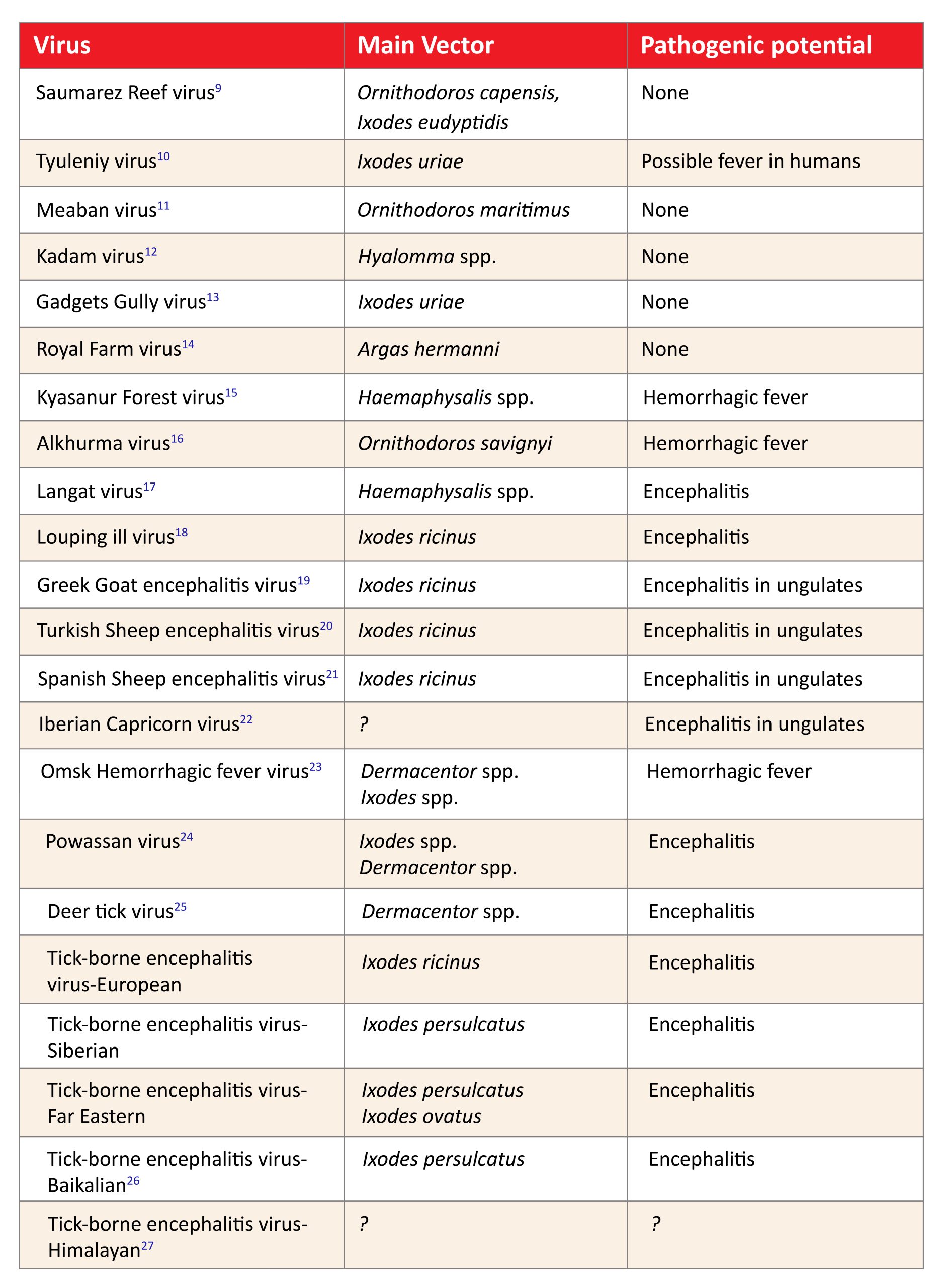
Click the image above to enlarge
The phylogeny and biogeography of tick-borne encephalitis virus
The genetic stability of tick-borne encephalitis virus
TBEV is a single stranded RNA virus, depending on a viral RNA-dependent RNA polymerase for its replication in cells. This kind of polymerase is known to be highly error prone due to the missing proof-reading mechanism.28 Therefore, RNA viruses exhibit a high mutation rate during replication leading to a phenomenon that almost any virus of a virus population has its own specific nucleotide sequence. Therefore, RNA virus populations are often named as “quasi-species”.29 In fact this high mutation rate in RNA viruses is thought to be one cause for species differentiation and adaption to new vectors or hosts.30 Therefore, the question arises, if the genetic information of TBE viruses, subtypes and strains is stable enough to calculate evolutionary traits and molecular clocks.
However, TBEV seems to be an exception of unstable RNA viruses. Several studies show that TBEV is genetically an extremely stable virus with only low variation. An analysis of strains from the Finish Island of Åland showed that even 44 years after first detection, TBEV strains are genetically almost identical and clearly distinguishable from other strains.31 Also, in non-published studies on the genetic variability of TBEV in a natural focus in Zillertal, Austria, the genetic variability of two strains that had been isolated 35 years apart from each other have a lower genetic variability than several isolates of the focus isolated during one single year (Dobler, unpublished observation). All data indicate that the TBEV develops a strain specificity in a particular TBEV focus and that this genetic specificity seems to remain stable for decades. This stability might be in part due to the assumed low replication rate in the vector and due to the fact that the virus has to pass two genetic bottlenecks: the vector and the natural host, and it May be that only virus strains optimally adapted to both biological systems are selected and may be successful in the natural transmission cycle. 32,33
The development of tick-borne encephalitis subtypes
“Tick-borne encephalitis virus” is the conventional name for the newly created virus species Orthoflavivirus encephalitidis, according to the new taxonomy of the International Committee on Taxonomy of Viruses (ICTV).2 This new taxonomic status does not distinguish between sub-species, subtypes, genotypes or serotypes. Therefore, the official status of the various subtypes of TBE virus in unclear. According to the commonly accepted notion based on earlier genetic analyses, TBEV strains differ in a subtype with a maximum of 2.5% on amino acid level and differences between TBEV virus subtypes is at about 5% at amino acid level.34 According to this classification a total of three subtypes could be identified at the beginning of genetic analyses of TBE virus.34
During the last 20 years a number of additional strains of TBEV were isolated and sequenced and at least three additional subtypes emerged using the criteria for TBEV subtypes, a Baikalian subtype, a Himalayan subtype and a subtype which has far only been identified once (strain 178/179).26,27,35,36 A newly detected TBE virus strain in The Netherlands, TBEV Sallandse, has an amino acid difference of >2% against all TBE virus strains from the
European subtype.37 The subtyping of this strain is still unclear. It might however lead in future to a second European subtype in the future.
The classification of Louping Ill (LI) virus is far from clear. Genetically it is closely related to the TBE-EU virus subtype. While in an earlier study TBEV and LI virus were genetically classified as only one species,3 a recent study confirmed a paraphyletic evolutionary development of TBEV and LI virus into two different species and proposed also two different species within the LI virus group, a Greek-Turkish species and a British-Spanish species of LI virus.38
For several years phylogenetic analyses were used to understand the differentiation of TBEV subtypes and their spread and distribution in Europe and in Asia. A first universal genetic calculation indicated that a common ancestor of the TBEV arrived some 3,000 years ago (range 1,700-4,800 years) ago in the area of Siberia. There it divided into two branches, one westward migrating genetic clade, finally forming the TBE-EU virus and LI virus and one eastward-migrating genetic clade which finally formed the TBE-Sib, TBE-Baik and TBE-FE virus subtypes. The TBE-Sib virus clade divided from the TBE-FE/-Baik clade about 2,500 years ago. The TBE-Baik clade and the TBE-178/79 virus diverged 1,400-1,800 years ago from the TBE-FE virus branch.6 According to these molecular calculations the ancestor of TBE-EU/LI viruses in Europe separated about 2,400 years ago into the Greek-Turkish clade of LI virus and about 1,700 years ago into the TBE-EU clade and LI-Spanish/-British clade.6 The TBE-EU virus strains as known today evolved only within the last 300 years (range 170-450 years) within Europe and spread there over large areas of the European continent.6 An earlier analysis of Czechoslovakian and German TBEV-EU subtype strains resulted in a geographic dispersal from Czechoslovakia to southeastern Germany around 50 years ago.39 The currently known geographic distribution of TBEV with its respective subtypes is shown in Figure 3.
Figure 3: Current knowledge of geographical distribution of TBE virus subtypes
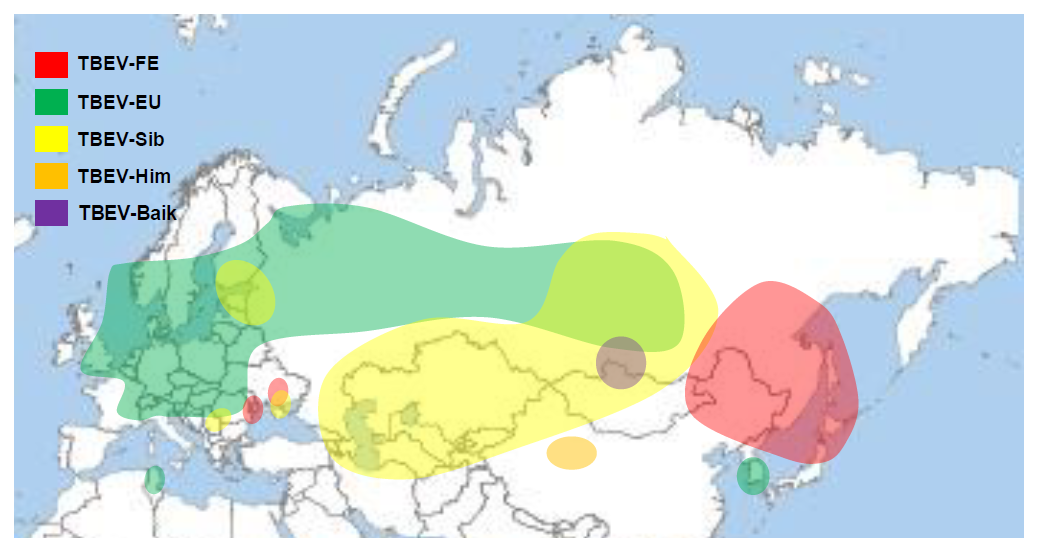
Click the image above to enlarge
The respective analyses on the genetic subtyping of the TBEV subtypes and their geographic association is still inconclusive for many of the strains. Best results are at the moment available for the Siberian subtype. A comprehensive genetic analysis of available TBE-Sib virus sequences can now clearly distinguish five genetic lineages or clades,40 of which
- one forms the Baltic lineage, one a single Serbian isolate,
- one isolates from the Kemerovo region in Western Siberia,
- one a large group of TBE-Sib viruses genetically related to the strain Vasilchenko and
- one related to the TBE-Sib virus strain Zausaev40
Similarly, clear genetic lineages cannot be identified in the currently available sequences of TBE-EU virus strains and TBE-FE virus strains. A comprehensive analysis of TBE-EU virus strains on the occasion of the phylogenetic classification of the TBE-EU virus strain “Sallandse”, shows at least 9 different genetic clades.37 Only one lineage shows geographic consistency and contains TBE-EU virus strains originating in Scandinavia and the Baltics. Otherwise, the genetic lineages are geographically mixed up and we have to discuss a panmictic genetic population structure of TBE-EU virus throughout Europe. Also, no conclusive data are yet available for the TBE-FE virus subtype. A recent analysis containing mainly Chinese genomes and sequences of TBE-FE virus subtype strains may imply the presence of potentially five genetic lineages or subclades of the TBE-FE virus subtype.41
Figure 4: Schematic phylogenetic tree of relevant human pathogenic flaviviruses
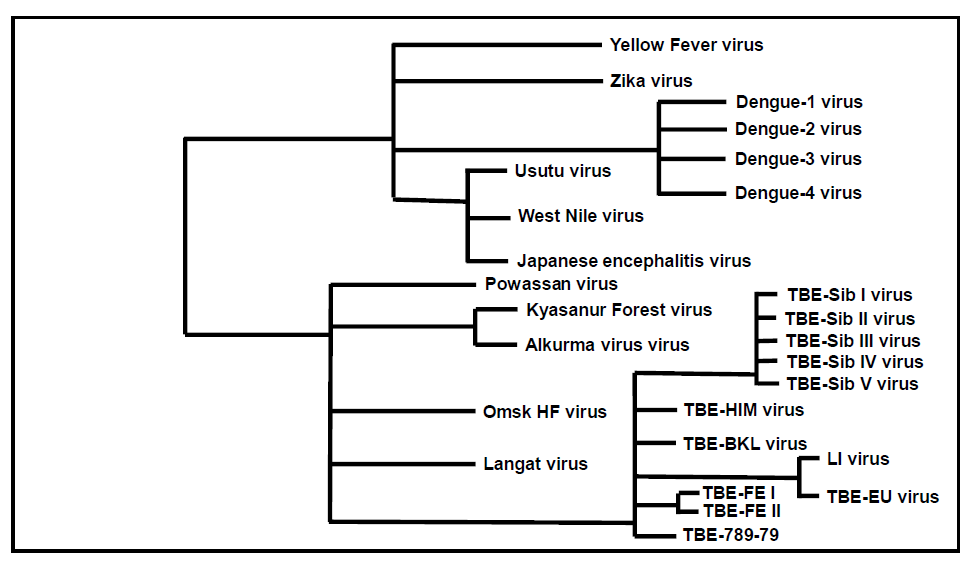
Click the image above to enlarge
Current ideas on the spread and dispersal of tick-borne encephalitis virus
The current knowledge on the geographic distribution and spread of TBEV subtypes raises a lot of questions. How could the TBE-EU emerge, as far as is known today, only in South Korea?42 At the Chinese-North Korean border region, the limited available data show only TBE-FE subtype strains.43 Similar questions arise for the TBE-FE virus strains detected in Ukraine and Moldova.44,45 The TBE-Sib virus identified in Serbia was shown to be a specific genetic lineage of the Siberian subtype most closely related to a Western Siberian genetic lineage of the TBE-Sib subtype viruses.40 Also the recent appearance of TBE-EU virus strains in Tunisia is still mysterious.46 At least some ideas about the occurrence of TBE-Sib virus in the Baltics and in Finland are currently discussed. One study showed an association with the construction of the Trans-Siberian Railway and the Trans-Siberian Highway.47 However recent studies, including genetic data from more strains which were isolated in the meantime, argue against these results and also speak of a “panmictic” biogeographic distribution of TBE-Sib virus strains in the Baltics and in Russia.48
These confusing data induce the general discussion on the ways TBEV spreads. In fact, only few studies have been conducted to date that elucidate the spread of TBEV. What we can clearly see is that we have to differentiate between a long-distance and short-distance spread of TBE virus. For long-distance spread, especially bird migration is a candidate for TBEV dispersal. Unfortunately, no studies are available to correlate genetic data of TBEV strains with bird migration routes. However, several studies show that birds may carry TBEV-infected ticks and therefore might be able to disperse them and other tick-borne flaviviruses during their migration.49,55 Earlier experiments on the potential of birds to become infected with TBEV and to keep high viral titers during migration indicate that birds may serve as hosts for newly blood-sucking ticks in other locations; however data remain inconclusive. 50,51,56-58, Nevertheless, there is some indirect evidence from phylogenetic studies of TBEV that birds may play a role in the spread of virus, e.g. on islands like England, Japan, the Finish Archipelago or also on the European mainland (e.g. Hungary).59-62
Beside long-distance spread TBEV also may spread on short distances from one microfocus to a new location nearby and thus form new microfoci. Again, this phenomenon has not been thoroughly studied so far. However, recent analyses on TBEV neighbouring natural foci showed that closely related TBEV strains can be isolated in nearby locations forming new microfoci.63 Although alternatively, dispersal of infected ticks by local birds cannot be excluded, the speculation may be allowed that also terrestrial game and other wild animals may transport and spread TBEV-infected ticks on a continuous way from one location to another.
Concluding remarks
Tick-borne flaviviruses are one of the medically most important groups of viruses. In sharp contrast to their huge medical importance, only little information is available on their origin, their evolution and also on their ways of spreading and distribution. Presented here are current data on the distribution of tick-borne flaviviruses with focus on the TBEV. Together with details on geographic distribution combined with phylogenetic information and results of spreading in the past, these concepts could lead to new concepts about the emergence of tick-borne orthoflaviviruses in the future.
Contact
Gerhard Dobler
gerharddobler@bundeswehr.org
Author
Citation
Dobler G. Tick-borne flavivirus complex – phylogeny and biogeography. Chapter 2. In: Dobler G, Erber W, Bröker M, Chitimia-Dobler L, Schmitt HJ, eds. The TBE Book. 7th ed. Singapore: Global Health Press; 2024. doi:10.33442/26613980_2-7
References
- Simmonds P, Becher P, Bukh J, et al. ICTV Virus Taxonomy Profile: Flaviviridae. J Gen Virol. 2017;98(1):2-3. doi:10.1099/jgv.0.000672.
- Postler TS, Beer M, Blitvich BJ, et al. Renaming of the genus Flavivirus to Orthoflavivirus and extension of binomial species names within the family Flaviviridae. Arch Virol. 2023;168(9):224. Published 2023 Aug 10. doi:10.1007/s00705-023-05835-1.
- Grard G, Moureau G, Charrel RN, et al. Genetic characterization of tick-borne flaviviruses: new insights into evolution, pathogenetic determinants and taxonomy. Virology. 2007;361(1):80-92. doi:10.1016/j.virol.2006.09.015.
- Pettersson JH, Fiz-Palacios O. Dating the origin of the genus Flavivirus in the light of Beringian biogeography. J Gen Virol. 2014;95(Pt 9):1969-1982.doi:10.1099/vir.0.065227-0.
- Moureau G, Cook S, Lemey P, et al. New insights into flavivirus evolution, taxonomy and biogeographic history, extended by analysis of canonical and alternative coding sequences. PLoS One. 2015;10(2):e0117849. Published 2015 Feb 26. doi:10.1371/journal.pone.0117849.
- Heinze DM, Gould EA, Forrester NL. Revisiting the clinal concept of evolution and dispersal for the tick-borne flaviviruses by using phylogenetic and biogeographic analyses. J Virol. 2012;86(16):8663-8671. doi:10.1128/JVI.01013-12.
- Zanotto PM, Gao GF, Gritsun T, et al. An arbovirus cline across the northern hemisphere. Virology. 1995;210(1):152-159. doi:10.1006/viro.1995.1326.
- Vogels CBF, Brackney DE, Dupuis AP 2nd, et al. Phylogeographic reconstruction of the emergence and spread of Powassan virus in the northeastern United States. Proc Natl Acad Sci U S A. 2023;120(16):e2218012120. doi:10.1073/pnas.2218012120.
- St George TD, Standfast HA, Doherty RL, Carley JG, Fillipich C, Brandsma J. The isolation of Saumarez Reef virus, a new flavivirus, from bird ticks Ornithodoros capensis and Ixodes eudyptidis in Australia. Aust J Exp Biol Med Sci. 1977;55(5):493-499. doi:10.1038/icb.1977.49.
- Chastel, C., et al., [Arbovirus serological survey among marine and non-marine birds of Brittany]. Bull Soc Pathol Exot Filiales. 1985;78(5):594-605.
- Chastel C, Main AJ, Guiguen C, et al. The isolation of Meaban virus, a new Flavivirus from the seabird tick Ornithodoros (Alectorobius) maritimus in France. Arch Virol. 1985;83(3-4):129-140. doi:10.1007/BF01309911.
- Burt FJ, Spencer DC, Leman PA, Patterson B, Swanepoel R. Investigation of tick-borne viruses as pathogens of humans in South Africa and evidence of Dugbe virus infection in a patient with prolonged thrombocytopenia. Epidemiol Infect.1996;116(3):353-361. doi:10.1017/s0950268800052687.
- St George TD, Doherty RL, Carley JG, et al. The isolation of arboviruses including a new flavivirus and a new Bunyavirus from Ixodes (Ceratixodes) uriae (Ixodoidea: Ixodidae) collected at Macquarie Island, Australia, 1975-1979. Am J Trop Med Hyg. 1985;34(2):406-412. doi:10.4269/ajtmh.1985.34.406.
- Williams RE, Casals J, Moussa MI, Hoogstraal H. Royal farm virus: a new tickborne group B agent related to the RSSE complex. Am J Trop Med Hyg. 1972;21(5):582-586. doi:10.4269/ajtmh.1972.21.582.
- Webb HE. Kyasanur Forest disease virus in three species on rodents. Trans R Soc Trop Med Hyg. 1965;59:205-211. doi:10.1016/0035-9203(65)90082-9.
- Dodd KA, Bird BH, Khristova ML, et al. Ancient ancestry of KFDV and AHFV revealed by complete genome analyses of viruses isolated from ticks and mammalian hosts. PLoS Negl Trop Dis. 2011;5(10):e1352. doi:10.1371/journal.pntd.0001352.
- Price WH, O’Leary W, Lee R, Parks J, Ganaway J. Studies of the virulence of langat virus propagated in chick embryo or hamster kidney tissue cultures. Am J Trop Med Hyg. 1963;12:782-786. doi:10.4269/ajtmh.1963.12.782.
- Casals J, Webster LT. Relationship of the Virus of Louping Ill in Sheep and the Virus of Russian Spring-Summer Encephalitis in Man. J Exp Med. 1944;79(1):45-63. doi:10.1084/jem.79.1.45.
- Papa A, Pavlidou V, Antoniadis A. Greek goat encephalitis virus strain isolated from Ixodes ricinus, Greece. Emerg Infect Dis. 2008;14(2):330-332. doi:10.3201/eid1402.070889.
- Whitby JE, Whitby SN, Jennings AD, Stephenson JR, Barrett AD. Nucleotide sequence of the envelope protein of a Turkish isolate of tick-borne encephalitis (TBE) virus is distinct from other viruses of the TBE virus complex. J Gen Virol. 1993;74 ( Pt 5):921-924. doi:10.1099/0022-1317-74-5-921.
- Marin MS, McKenzie J, Gao GF, Reid HW, Antoniadis A, Gould EA. The virus causing encephalomyelitis in sheep in Spain: a new member of the tick-borne encephalitis group. Res Vet Sci. 1995;58(1):11-13. doi:10.1016/0034-5288(95)90081-0.
- Mansfield KL, Morales AB, Johnson N, et al. Identification and characterization of a novel tick-borne flavivirus subtype in goats (Capra hircus) in Spain. J Gen Virol. 2015;96(Pt 7):1676-1681. doi:10.1099/vir.0.000096.
- Růžek D, Yakimenko VV, Karan LS, Tkachev SE. Omsk haemorrhagic fever. Lancet. 2010;376(9758):2104-2113. doi:10.1016/S0140-6736(10)61120-8.
- CASALS J. Antigenic relationship between Powassan and Russian spring-summer encephalitis viruses. Can Med Assoc J. 1960;82(7):355-358.
- Solomon IH, Spera KM, Ryan SL, et al. Fatal Powassan Encephalitis (Deer Tick Virus, Lineage II) in a Patient With Fever and Orchitis Receiving Rituximab. JAMA Neurol. 2018;75(6):746-750. doi:10.1001/jamaneurol.2018.0132.
- Sukhorukov GA, Paramonov AI, Lisak OV, et al. The Baikal subtype of tick-borne encephalitis virus is evident of recombination between Siberian and Far-Eastern subtypes. PLoS Negl Trop Dis. 2023;17(3):e0011141.
- Dai X, Shang G, Lu S, Yang J, Xu J. A new subtype of eastern tick-borne encephalitis virus discovered in Qinghai-Tibet Plateau, China. Emerg Microbes Infect. 2018;7(1):74. Published 2018 Apr 25. doi:10.1038/s41426-018-0081-6.
- Steinhauer DA, Domingo E, Holland JJ. Lack of evidence for proofreading mechanisms associated with an RNA virus polymerase. Gene. 1992;122(2):281-288. doi:10.1016/0378-1119(92)90216-c.
- Asghar N, Pettersson JH, Dinnetz P, Andreassen Å, Johansson M. Deep sequencing analysis of tick-borne encephalitis virus from questing ticks at natural foci reveals similarities between quasispecies pools of the virus. J Gen Virol. 2017;98(3):413-421. doi:10.1099/jgv.0.000704.
- Helmová R, Hönig V, Tykalová H, Palus M, Bell-Sakyi L, Grubhoffer L. Tick-Borne Encephalitis Virus Adaptation in Different Host Environments and Existence of Quasispecies. Viruses. 2020;12(8):902. Published 2020 Aug 18. doi:10.3390/v12080902.
- Gritsun TS, Lashkevich VA, Gould EA. Tick-borne encephalitis. Antiviral Res. 2003;57(1-2):129-146. doi:10.1016/s0166-3542(02)00206-1.
- Romanova LIu, Gmyl AP, Dzhivanian TI, et al. Microevolution of tick-borne encephalitis virus in course of host alternation. Virology. 2007;362(1):75-84. doi:10.1016/j.virol.2006.12.013.
- Ecker M, Allison SL, Meixner T, Heinz FX. Sequence analysis and genetic classification of tick-borne encephalitis viruses from Europe and Asia. J Gen Virol. 1999;80 ( Pt 1):179-185. doi:10.1099/0022-1317-80-1-179.
- Kozlova, I.V., et al., Genetic and Biological Properties of Original TBEV Strains Group Circulating in Eastern Siberia, in Encephalitis. 2013.
- Adelshin RV, Sidorova EA, Bondaryuk AN, et al. “886-84-like” tick-borne encephalitis virus strains: Intraspecific status elucidated by comparative genomics. Ticks Tick Borne Dis. 2019;10(5):1168-1172. doi:10.1016/j.ttbdis.2019.06.006.
- Hoornweg TE, Godeke GJ, Hoogerwerf MN, et al. Rescue and in vitro characterization of a divergent TBEV-Eu strain from the Netherlands. Sci Rep. 2023;13(1):2872. Published 2023 Feb 18. doi:10.1038/s41598-023-29075-0.
- Bondaryuk AN, Andaev EI, Dzhioev YP, et al. Delimitation of the tick-borne flaviviruses. Resolving the tick-borne encephalitis virus and louping-ill virus paraphyletic taxa. Mol Phylogenet Evol. 2022;169:107411. doi:10.1016/j.ympev.2022.107411.
- Weidmann M, Růžek D, Křivanec K, et al. Relation of genetic phylogeny and geographical distance of tick-borne encephalitis virus in central Europe. J Gen Virol. 2011;92(Pt 8):1906-1916. doi:10.1099/vir.0.032417-0.
- Tkachev SE, Babkin IV, Chicherina GS, et al. Genetic diversity and geographical distribution of the Siberian subtype of the tick-borne encephalitis virus. Ticks Tick Borne Dis. 2020;11(2):101327. doi:10.1016/j.ttbdis.2019.101327.
- Li X, Ji H, Wang D, et al. Molecular detection and phylogenetic analysis of tick-borne encephalitis virus in ticks in northeastern China. J Med Virol. 2022;94(2):507-513. doi:10.1002/jmv.27303.
- Kim SY, Jeong YE, Yun SM, Lee IY, Han MG, Ju YR. Molecular evidence for tick-borne encephalitis virus in ticks in South Korea. Med Vet Entomol. 2009;23(1):15-20. doi:10.1111/j.1365-2915.2008.00755.x.
- He X, Zhao J, Fu S, et al. Complete Genomic Characterization of Three Tick-Borne Encephalitis Viruses Detected Along the China-North Korea Border, 2011. Vector Borne Zoonotic Dis. 2018;18(10):554-559. doi:10.1089/vbz.2017.2173.
- Ponomareva EP, Mikryukova TP, Gori AV, et al. Detection of Far-Eastern subtype of tick-borne encephalitis viral RNA in ticks collected in the Republic of Moldova. J Vector Borne Dis. 2015;52(4):334-336.
- Yurchenko OO, Dubina DO, Vynograd NO, Gonzalez JP. Partial Characterization of Tick-Borne Encephalitis Virus Isolates from Ticks of Southern Ukraine. Vector Borne Zoonotic Dis. 2017;17(8):550-557. doi:10.1089/vbz.2016.2094.
- Fares W, Dachraoui K, Cherni S, et al. Tick-borne encephalitis virus in Ixodes ricinus (Acari: Ixodidae) ticks, Tunisia. Ticks Tick Borne Dis. 2021;12(1):101606. doi:10.1016/j.ttbdis.2020.101606.
- Kovalev SY, Chernykh DN, Kokorev VS, Snitkovskaya TE, Romanenko VV. Origin and distribution of tick-borne encephalitis virus strains of the Siberian subtype in the Middle Urals, the north-west of Russia and the Baltic countries. J Gen Virol. 2009;90(Pt 12):2884-2892. doi:10.1099/vir.0.012419-0.
- Deviatkin AA, Kholodilov IS, Belova OA, et al. Baltic Group Tick-Borne Encephalitis Virus Phylogeography: Systemic Inconsistency Pattern between Genetic and Geographic Distances. Microorganisms. 2020;8(10):1589. Published 2020 Oct 15. doi:10.3390/microorganisms8101589.
- Capligina V, Salmane I, Keišs O, et al. Prevalence of tick-borne pathogens in ticks collected from migratory birds in Latvia. Ticks Tick Borne Dis. 2014;5(1):75-81. doi:10.1016/j.ttbdis.2013.08.007.
- Ernek E, Kozuch O, Lichard M, Nosek J. The role of birds in the circulation of tick-borne encephalitis virus in the Tribec region. Acta Virol. 1968;12(5):468-470.
- Gresikova M, Nosek J, Rehacek J, Albrecht P. The role of birds in a natural focus of tick-borne encephalitis. II. Experimental infection of great tits (Parus major L.) with tick-borne encephalitis virus. J Hyg Epidemiol Microbiol Immunol. 1962;6:339-342.
- Hoffman T, Lindeborg M, Barboutis C, et al. Alkhurma Hemorrhagic Fever Virus RNA in Hyalomma rufipes Ticks Infesting Migratory Birds, Europe and Asia Minor. Emerg Infect Dis. 2018;24(5):879-882. doi:10.3201/eid2405.171369.
- Likar M, Laneger M, Pintar G, Belcijan I. Infections with arthropod borne viruses in wild birds in an area with endemic central European tick borne encephalitis. Pathol Microbiol (Basel). 1963;26:285-297. doi:10.1159/000161377.
- Wilhelmsson P, Jaenson TGT, Olsen B, Waldenström J,Lindgren PE. Migratory birds as disseminators of ticks and the tick-borne pathogens Borrelia bacteria and tick-borne encephalitis (TBE) virus: a seasonal study at Ottenby Bird Observatory in South-eastern Sweden. Parasit Vectors.2020;13(1):607. Published 2020 Dec 3. doi:10.1186/s13071-020-04493-5.
- Waldenström J, Lundkvist A, Falk KI, et al. Migrating birds and tickborne encephalitis virus. Emerg Infect Dis. 2007;13(8):1215-1218. doi:10.3201/eid1308.061416.
- Sobeslavsky, O., F. Rehn, and J. Fischer, [Isolation oftickborne encephalitis virus from moor hen (Fulica atra)].Cesk Epidemiol Mikrobiol Imunol, 1960;9:256-61.
- Dumina, A.L. and E.S. Sarmanova, [Investigations on virusemia in wild animals and birds experimentally infected with tick-borne vernal encephalitis virus]. Vopr Virusol. 1956;1(3):23-6.
- Gresíková M, Sekeyová M, Prazniaková E. Isolation and identification of group B arboviruses from the blood of birds captured in Czechoslovakia. Acta Virol. 1975;19(2):162-164.
- Holding M, Dowall S, Hewson R. Detection of tick-borne encephalitis virus in the UK. Lancet. 2020;395(10222):411.doi:10.1016/S0140-6736(20)30040-4.
- Kentaro Y, Yamazaki S, Mottate K, et al. Genetic and biological characterization of tick-borne encephalitis virus isolated from wild rodents in southern Hokkaido, Japan in 2008. Vector Borne Zoonotic Dis. 2013;13(6):406-414. doi:10.1089/vbz.2012.1231.
- Smura T, Tonteri E, Jääskeläinen A, et al. Recent establishment of tick-borne encephalitis foci with distinct viral lineages in the Helsinki area, Finland. Emerg Microbes Infect. 2019;8(1):675-683.doi:10.1080/22221751.2019.1612279.
- Egyed L, Rónai Z, Dán Á. Hungarian tick-borne encephalitis viruses isolated from a 0.5-ha focus are closely related to Finnish strains. Ticks Tick Borne Dis. 2018;9(5):1064-1068. doi:10.1016/j.ttbdis.2018.03.032.
- Lang D, Chitimia-Dobler L, Bestehorn-Willmann M, et al. The Emergence and Dynamics of Tick-Borne Encephalitis Virus in a New Endemic Region in Southern Germany. Microorganisms. 2022;10(11):2125. Published 2022 Oct 27. doi:10.3390/microorganisms10112125.