Chapter 10:
TBE in animals
Martin Pfeffer, Hannah M. Schmuck, Michael Leschnik
Key points
- TBEV-infection can cause symptomatic disease in dogs and horses, similar to the TBE in humans.
- Microbiological confirmation of TBEV infection in animals is similar to diagnostics in humans.
- Domestic ruminants may serve source of human infection via the alimentary routs (dairy products).
- Small mammals play the major role as the reservoir for the TBEV and are thus of utmost epidemiological relevance.
- Other species like cervids and wild boar are of interest for sentinel surveillance, as their seropositivity in a specific region indicates the presence of a natural TBEV-focus.
Introduction
While tick-borne encephalitis (TBE) is well documented as a public health threat, the veterinary aspects of this zoonotic disease are little recognized. TBE in animals has, for very long, been considered to be a problem exclusive to domestic ruminants due to their known potential to transmit tick-borne encephalitis virus (TBEV) via raw milk and raw milk products to consumers. While clusters of such cases continuously declined with the invention of milk pasteurization and overall improvements in hygiene management in cattle farming, goats and sheep flocks are still kept in traditional grazing farms where they are exposed to TBEV-infected ticks.1,2 In other words, even in industrialized countries, consumption of raw milk products continues to be a risk factor to acquire a TBEV infection. As society continues to exhibit a trend towards a preference for “natural products” (assuming consumers can afford these), alimentary TBEV infections may be observed more frequently in the future. While this is a ‘direct’ zoonotic aspect of TBE (besides the tick bite of course), animals play a role in TBEV transmission in many other ways; either as diseased dead-end hosts, as infected animals without obvious burden of disease, or in maintaining and spreading the virus itself.
Dogs
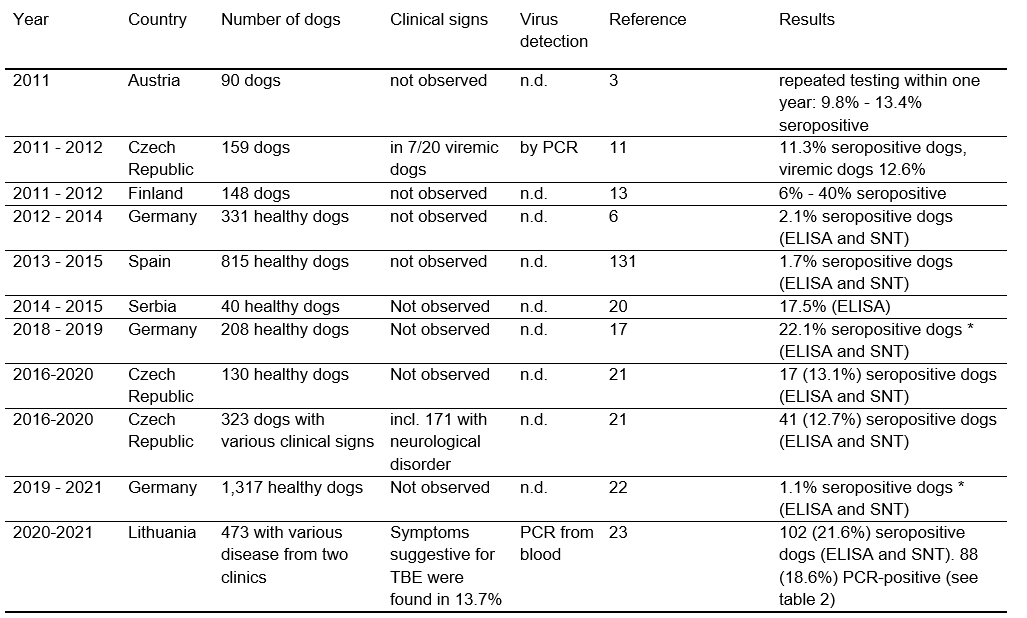
Canine TBEV infection is a frequent event in endemic areas, with a calculated annual risk of about 11.6%.3 Total seroprevalence in the canine population has been examined in several countries: Switzerland 3.6–5.9%,4 Greece 1–8%,5 Germany 2.1– 42.7%,6,7 Belgium 0.1%,8 Denmark 4.8–30%,9 Czech Republic 3.3–11.3%,10,11 Norway 16.4%,12 Finland 6–40%,13 and Austria 13.3–24%.3,14 Since inclusion criteria were different regarding the presence of clinical symptoms, residence, and tick-exposure of the examined dogs, results are difficult to compare (Table 1). Different test systems (enzyme-linked immunosorbent assay [ELISA], serum neutralization test [SNT]) used in these studies clearly influenced the results too. TBE has always been stated to be a tick-borne infection, mainly transmitted by ticks of the genus Ixodes; however, Dermacentor reticulatus ticks may play an important role in transmission to dogs.15 There has been one single case of a dog from the Czech Republic with a TBE-infection suspected to be due to consumption of raw goat milk.10 Regardless of the way dogs get infected, a recent study showed that walking a dog is a risk factor for human infections.16
| Year | Number of Cases | Incidence / 105 |
|---|---|---|
| 1995 | 5 | 0.10 |
| 1996 | 8 | 0.16 |
| 1997 | 19 | 0.38 |
| 1998 | 16 | 0.31 |
| 1999 | 12 | 0.23 |
| 2000 | 42 | 0.81 |
| 2001 | 33 | 0.64 |
| 2002 | 38 | 0.73 |
| 2003 | 16 | 0.31 |
| 2004 | 29 | 0.56 |
| 2005 | 16 | 0.31 |
| 2006 | 18 | 0.34 |
| 2007 | 20 | 0.38 |
| 2008 | 23 | 0.43 |
| 2009 | 25 | 0.47 |
| 2010 | 38 | 0.71 |
| 2011 | 43 | 0.80 |
| 2012 | 39 | 0.72 |
| 2013 | 38 | 0.71 |
| 2014 | 47 | 0.86 |
| 2015 | 68 | 1.24 |
| 2016 | 61 | 1.11 |
| 2017 | 82 | 1.49 |
| 2018 | 79 | 1.43 |
| 2019 | 69 | 1.25 |
| 2020 | 91 | 1.64 |
| 2021 | 148 | 2.67 |
| 2022 | 124 | 2.23 |
Figure 1: A Rottweiler during recovery after chronic disease over 3 months – remarkable weight loss due to systemic muscle atrophy

Figure 2: Acute head pressing with concurrent compulsive walking and disorientation on day 2 of a dog with TBE
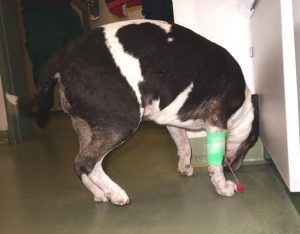
Figure 3: A male Spitz with central vestibular dysfunction and left-sided Horner syndrome during acute TBE
Course of TBE
Despite frequent TBEV infections in dogs, most of them do not develop any clinical signs.17 Dogs seem to be less susceptible than humans, although a lethal outcome within the first week of disease is documented in 16–50% of clinically symptomatic cases. Infection may lead to an acute course of the disease, with complete remission of symptoms within 1–2 weeks (31–59%). Infrequently, prolonged disease courses are described with long time period to remission (12–25%). These dogs frequently suffer from late sequela–like paresis, muscle atrophy, epileptic seizures, or blindness (Figure 1).10,18,19,27,28
Clinical pictures
After an estimated incubation period of 5–9 days, first clinical symptoms occur and develop to a maximum level within 48 hours. Initially, most dogs are depressed and show non-specific signs such as salivation and vomiting (25%), refusal to eat, and are reluctant to move due to generalized weakness, although some dogs show compulsive walking, circling to one side (25%), unusual behavior (70–91%), and head pressing (Figure 2).10,27-30
The elevated body temperature (42–66%) may initially be classed as fever; later on, it is more likely a result of non-voluntary excessive muscle contraction (e.g., seizures, loss of inhibition by upper motor neuron damage). Seizures are a principal result of cerebral damage due to TBEV infection and are observed in 12–33% of canine cases.28,30 Neurological symptoms like paresis (8–38%), vocalization due to painful perception of active and passive back movement (21–66%), and deficits of the cranial nerves (16–50%) (Figure 3) develop within a few hours thereafter.28,30,31
Blindness due to papillitis, optic nerve inflammation, or chiasma opticus neuritis may become the dominant symptom and systemic signs may diminish. Visual deficits may be the major clinical sign of disease and result from detachment of peripapillary retina, peripapillary hemorrhages, and inflammatory edema.32,33 Degeneration and demyelination of cranial nerves is certainly initiated by the virus’ neurotropism. Later on, secondary immune reaction to neural tissue may prolong the period of damage and lead to irreversible symptoms such as retinal and optic disc atrophy. Other cranial nerve deficits like trigeminal dysfunction, resulting in reduced facial sensation and chewing muscle atrophy, vestibular signs (nystagmus and positional strabismus, Figure 4), and facial palsy, are observed.
Figure 4: A comatose dog in lateral recumbency with severe brain stem encephalitis leading to anisocoria and left-sided strabismus

Major involvement of the spinal cord results in mostly symmetrical paresis, muscle twitching, and proprioceptive dysfunction (38-50%), which may also be present as an exclusive symptom and may occur asymmetrically (Figure 5).10,28,30,31
Figure 5: A case of canine TBE with hemiparesis and spontaneous dorsal paw placement

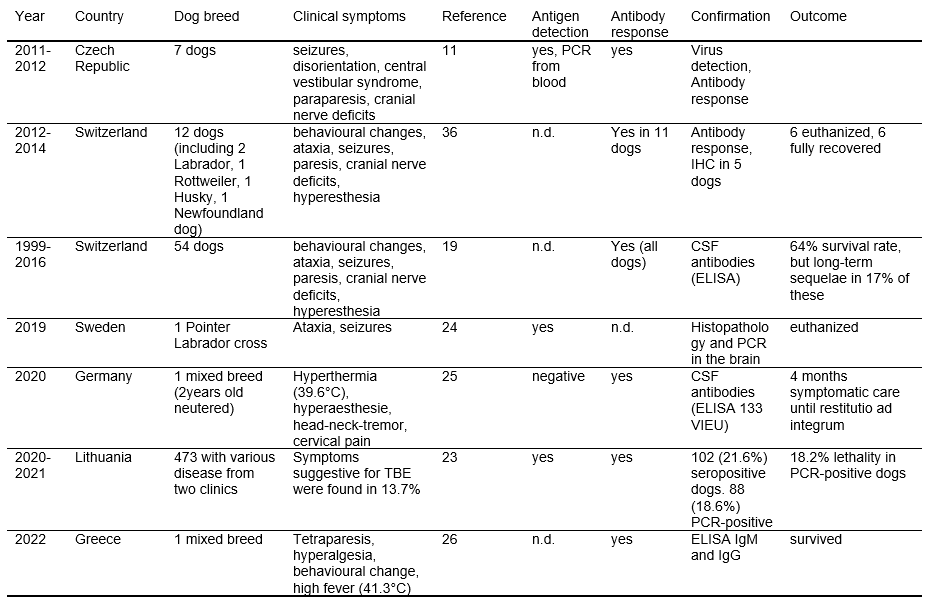
There is no significant breed, gender, or age predisposition, although most cases are described in adult middle- to large-breed dogs. Rottweilers and Huskies are overrepresented in the literature14,31,32 (Table 2).
Brainstem symptoms like arrhythmical breathing pattern may be present in comatose dogs, especially in severe cases with guarded prognosis.
Video: Comatose dog of Figure 4 with arhythmical breathing indicative of brain stem lesion
Involvement of the brainstem may result in symptoms like arrhythmical breathing and disorder of other vital functions. Prognosis of such severe cases is very guarded. Major involvement of the spinal cord results in mostly symmetrical paresis, muscle twitching, and proprioceptive dysfunction (38-50%), which may also be present as an exclusive symptom and may occur asymmetrically (Figure 5).10,28,30,31
There is no significant breed, gender, or age predisposition, although most cases are described in adult middle- to large-breed dogs. Rottweilers and Huskies are over-represented in the literature14,31,32 (Table 2).
| Year | Number of Cases | Incidence / 105 |
|---|---|---|
| 1995 | 5 | 0.10 |
| 1996 | 8 | 0.16 |
| 1997 | 19 | 0.38 |
| 1998 | 16 | 0.31 |
| 1999 | 12 | 0.23 |
| 2000 | 42 | 0.81 |
| 2001 | 33 | 0.64 |
| 2002 | 38 | 0.73 |
| 2003 | 16 | 0.31 |
| 2004 | 29 | 0.56 |
| 2005 | 16 | 0.31 |
| 2006 | 18 | 0.34 |
| 2007 | 20 | 0.38 |
| 2008 | 23 | 0.43 |
| 2009 | 25 | 0.47 |
| 2010 | 38 | 0.71 |
| 2011 | 43 | 0.80 |
| 2012 | 39 | 0.72 |
| 2013 | 38 | 0.71 |
| 2014 | 47 | 0.86 |
| 2015 | 68 | 1.24 |
| 2016 | 61 | 1.11 |
| 2017 | 82 | 1.49 |
| 2018 | 79 | 1.43 |
| 2019 | 69 | 1.25 |
| 2020 | 91 | 1.64 |
| 2021 | 148 | 2.67 |
| 2022 | 124 | 2.23 |
Laboratory findings and diagnosis
A definite diagnosis in dogs with TBE is rarely achieved intra vitam as it has to be supposed to be very unlikely to detect the virus in the blood or in the cerebrospinal fluid (CSF). In one study from the Czech Republic, 12.6% of canine blood samples tested positive for TBEV by nested RT-polymerase chain reaction (PCR), although only one-third of these dogs suffered from neurological symptoms.11
Whether the other dogs were in an asymptomatic carrier status, or just happened to be tested during their viremic phase with uncharacteristic symptoms, as reported in humans, remained unclear. Virus detection in the CSF has been achieved only in single cases within the first 3 days of disease.30 Immunological rapid virus clearance in the dog’s brain and CSF seems to be very fast and completed before most diagnostic procedures are performed. The inability of the central nervous system’s (CNS) local immune system to eliminate the virus within a few days is probably the reason for a fatal outcome, as in most of these cases no specific intrathecal antibody production and no increased cell count in the CSF were detected prior to death.28 CSF analysis in affected dogs with clinical signs mostly reveals elevated leukocyte count, with predominantly mononuclear cells and elevated total protein. CSF changes are concomitant to virus elimination and rising antibody titers.
Specific antibodies are detectable in the serum of affected dogs within a few days.7,28,29,31 Comparison of a commercially available all species ELISA, indirect IFT and SNT using a panel of 208 dog sera revealed a sensitivity of 78.3% and 84.8% when compared to SNT and a specificity of 98.8% and 99.4%. IIFT an ELISA are thus good in case of confirming clinical cases with suspicion of TBE but due to the deficits in sensitivity the SNT is superior in epidemiological studies.17 As TBEV is a biosafety level 3 agent in many countries, production of the antigen used for any serological test is limited to facilities with an appropriate safety level. In order to circumvent this obstacle two assays have been developed using prME subviral particles expressed through a Semliki Forest Virus-based expression system. In one assay this antigen is used in a Vero cells system analogous to an IFT, while the other one is a capture ELISA using a monoclonal antibody (MAB1418) which specifically binds to domain III of glycoprotein E of TBEV. Specificity was thus raised to 100% for both assays making it suitable for epidemiological applications.33 For clinical confirmation the detection of positive CSF IgG antibodies is recommended.34 Cross-reactivity to Louping ill virus, West Nile virus, and Usutu virus should be taken into consideration in endemic areas.10,35 Magnetic resonance imaging findings included bilateral and symmetrical gray matter lesions involving the thalamus, hippocampus, brain stem, basal nuclei, and ventral horn on the spinal cord.
All lesions had minimal or no mass effect, or perilesional edema.36 These findings are comparable to the distribution of lesions in the canine brain detected by necropsy and immunohistochemistry.37
Proton magnetic resonance spectroscopy, to evaluate metabolic abnormalities in dogs with TBE, revealed significant differences with dogs with immune mediated meningoencephalitis and healthy dogs.38
A tentative diagnosis of TBE in dogs should fulfill the following criteria: tick exposure or observed tick infestation, neurological signs indicative for a diffuse or multifocal CNS disease, (mostly mononuclear) pleocytosis in the CSF, a positive antibody titer in serum or CSF, or in the case of fatal outcome a positive virus confirmation within the brain or spinal cord. In the future, highly sensitive PCR techniques may include virus detection in the diagnostic work-up in early stages of disease. Increasing serum titers may be detected, but more often rapidly decreasing titers are observed when dogs reach partial or complete remission of clinical signs.28,38
Possible differential diagnoses include other viral meningoencephalitis such as distemper, rabies, pseudorabies, as well as protozoal, bacterial, or fungal meningoencephalitis, and paraneoplastic and immune mediated meningoencephalitis.
Figure 6: An old Labrador Retriever during rehabilitation. Water training over months improved muscle strength and coordination

Treatment
Symptomatic therapy is strongly recommended for dogs with TBE. Water and food maintenance orally, by constant rate infusion, or by gastric tubes and supportive care is essential. Sedation and relaxation are necessary in the case of seizures. Steroid use is controversial, as immunosuppression may prolong the presence of the virus. In dogs with marked CSF pleocytosis, steroids seem to be mandatory to effectively protect the brain tissue from further fulminant immune response. In cases of muscle atrophy and paresis, physiotherapy (Figure 6) as early as possible has been shown to improve the general outcome and shorten the time of rehabilitation.30,31
Prevention
There is no licensed anti-TBE vaccine for dogs, although they develop detectable antibody titers after vaccination with a human vaccine.39 In a recent study dogs were infected either with 108 pfu or 106 pfu TBEV strain 9001 isolated from Ixodes ricinus ticks in the Czech Republic in 1978 (back then Czechoslovakia). All animals developed no overt clinical signs but high IgG antibody titers in ELISA and high SNT antibody titers demonstrating that dogs are capable of mounting protectable Immune response upon infection.21 With the new European Animal Health Law vaccines for animals are handled in the EU like other animal therapeutics. In case no licensed vaccine against TBE is available for dogs the so-called cascade can be used, which in this case means that human vaccine can be rededicated by the treating veterinarian upon request at the responsible veterinary authorities. Depending on the size (and weight) of the dog, the pediatric formulation is recommend. Recently, colleagues tested whole virus inactivated TBEV strain Hypr as vaccine for dogs and found it well tolerated and to elicit a protective immunity.132
Tick protection is the most important measure to avoid transmission and infection, mainly performed by regular administration of acaricidal substances (spot on, tablets, shampoos, collars) and immediate tick removal after detection by the owner.3 Regular anti-tick measures are essential to reduce transmission risk all through the year as single canine cases have been reported even during the cold seasons of the year.32
Horses
Although the first clinical case of laboratory-confirmed TBE in a horse was published more than 35 years ago,40 our knowledge about the impact of TBEV in horse populations is still scarce. There are only few published studies where clinical signs of neurological disorder could be traced to the TBEV as etiology. After the aforementioned initial published case from Switzerland, 8 horses with clinical symptoms were described in Austria, 2 of which were severely ill;41 1 out of 3 diseased animals from a study in Germany had to be euthanized;42 and again in Germany, some years later, an infected animal had to be euthanized.43 A case in Austria with the same outcome was worked up in a very thorough way, excellently describing the symptoms and laboratory finding. The authors in addition provide a video as supplementary to the manuscript which shows the 16 year-old horse with its neurological symptoms.44 The clinical picture in horses mirrors that which we described for dogs, displaying a broad spectrum of neurological symptoms: ataxia, tonic-clonic seizures, apathy and stupor, inappetence, mydriasis, convulsions of the legs, skittishness, bruxism, and altered reactions to environmental stimuli. Regarding therapeutic options and prognosis, a horse with recumbent status due to TBE has a poor prognosis as long as it is not possible to force the horse to stand up again.
The few case reports available suggest that clinical TBE in horses is a rare event, although basic horse population-based data are missing. Looking at the few seroprevalence studies in horses, the prevalence of anti-TBE-antibodies ranged from 26.1% and 13% in Austria43,48 to 2.9% in central Germany,42 0.8% in northern Germany,22 3.7% and 5.6% in eastern Germany,46,47 and 5.2% and 23.4% in southern Germany43,48 to 0 of 40 horses investigated in Hungary49 or 0 of 2349 horses from the Czech Republic.50 Even in Spain a seroprevalence of 3.1% was reported in horses.51 In Serbia and Croatia 5% and 12.2% of horses showed specific antibodies against TBEV.20,52 The highest prevalence in horses was reported in a cross-section study from Lithuania with 37.5% reflecting the high human incidence there. Remarkable in this study was that 3.9% of theses horses also had a viremia based on the detection of viral RNA by RT-PCR, but none of the horses showed any overt signs of sickness.53 Cross-reactivity to other flavivirus may influence these results.50,54 Horses have been suggested to be good sentinel animals for human TBEV infection risk, because they readily seroconvert upon infection, but they stick more to a given territory in comparison to dogs who, as family members, travel more.
Domestic ruminants
For more than half a century, grazing cattle, goats, and sheep have been known to be susceptible to TBEV infection. Interestingly, these ruminants do not develop any clinical symptoms, and even after experimental infection, a slight elevation of body temperature is a rare finding.55,56 However, in 2015, a five-month-old lamb in Bavaria displayed neurological symptoms , and after euthanasia, TBEV infection was diagnosed.57 Whether this case was the result of an unknown underlying disease or immunosuppressive factors cannot be determined. TBE in domestic ruminants, if it occurs at all, appears to be an extreme exception. Nevertheless, infected animals develop viremia with a duration of up to 19 days.58 A study in the Swiss canton of Valais found 4.25% of the tested goats to be seropositive according to an ELISA test, with 40.4% of these testing positive on a serum neutralization test (SNT).59 In the canton of Ticino, officially labelled as non-TBE-endemic, SNT-positive goats were found in 10 out 37 of herds (14.6% out of 662 sera).60 In Germany the intra-herd prevalence in sheep and goat herds was between 2.3% and 25%, but antibodies were found in some of the districts not considered TBE endemic area, thus reflecting the human situation and arguing for small ruminant as good sentinel animals for human infection risk.61 In Sweden serology from sheep milk was successfully used to map what they called “TBEV hotspots”.62
Even if viremia is shorter than 1 week, virus is shed via milk and remains infectious in cheese or other products prepared from unpasteurized milk. Consumption of such products may have led to an alimentary infection of a group of individuals who became infected through the same batch of contaminated food, resulting in clusters of human cases.63 Such clusters of cases have recently been reviewed2 and were thought to be restricted to nations in Eastern Europe with Slovakia having the highest occurrence of alimentary TBE outbreaks in Europe.64 But alimentary infections due to the consumption of raw milk products are also reported from countries with rather low tick-borne incidences, like Croatia.65,66 However, alimentary TBEV infection with clinical TBE occurred recently in Germany as a result of consumption of fresh raw goat milk.67 As there is a growing trend towards consumption of natural food products in the industrialized nations of Western Europe, such scenarios may be witnessed more frequently in the future. One study in an endemic region in Poland found TBEV in milk from sheep (22.2%), goats (14.8%), and cows (11.1%).68 In Norway, a study found TBEV RNA in 5.4% of tested raw milk samples. Positive blood serum samples only occurred in one municipality, where 88.2% of tested cows had specific antibodies. Remarkably, none of the cows with a positive milk sample had detectable antibodies and vice versa.69 Domestic ruminants do develop an antibody response, which in the case of goats and sheep is measurable for at least 28 months or even up to 6 years and 10 months.35,39,70 Exposure to TBEV seems not to result uniformly in seroconversion of the entire flock of animals.71,72 Whether this indicates that not all animals of the same herd were exposed and infected or that some animals did not mount an immune response is not known. Also, the extent of antibody response seems to vary between the species.73
Game animals (wild boar, cervids, foxes)
Roe deer (Capreolus capreolus) are the most abundant cervids in Germany, sharing their habitat with ticks everywhere. They are well known as hosts for nymphs and adult ticks and thus are as important to maintenance of the tick population as the small mammals are for larvae and nymphs (see below). It is common to find hundreds of ticks per individual and, consequently, the odds of roe deer becoming infected in TBE-endemic areas are rather high.74 Therefore, they can be a useful tool to identify endemic areas as could be seen in the Netherlands, where TBE was regarded as an imported disease until 2016. Serologic screening there showed TBEV-neutralizing antibodies with a seroprevalence of 2% in roe deer.75 Clinical or pathological signs that raise suspicions of an overt TBEV infection had never been described for roe deer until recently with a single case in Italy.76 Seroconversion after infection seems to be the rule, and this fact has been widely used to estimate TBE prevalence in certain areas. As roe deer are territorial animals, many researchers claim that this serological data could be very useful in finding and describing possible TBE-endemic areas, in particular in low-endemic areas or regions in which TBE cases in humans are reported only sporadically.77-84 The discrepancy of often double-digit percentages of seroprevalence in roe deer and no, or almost no, human cases is puzzling, and needs to be investigated further. As TBEV is known to be circulating in such areas, an understanding of why only few or no human cases occur could be key to developing strategies aimed at reducing TBE incidence in high-endemic areas (as defined by the number of human cases).
Likewise, the wild boar (Sus scrofa) is present all over Europe and is commonly infested with ticks. There are no records of a possible TBE-like disease in wild boar and only 2 studies investigated the seroprevalence against TBEV in wild boar. Nevertheless, these studies demonstrated a surprisingly high percentage of animals with antibodies against TBEV in areas with no notified human TBE cases.82 A sero-survey of wild boar in Belgium revealed the presence of TBEV, with 2.9% of the 238 wild boar investigated having specific neutralizing antibodies against TBEV.85 As Belgium is considered to be traditionally free of autochthonous TBE,2,86,87 this study demonstrates the power of using animal surveillance data for pinpointing TBE-endemic areas. A similar approach was applied in France using wild boar and roe deer sera with similar results, i.e. 2.9% and 0.3% seropositive animals.88 Like the roe deer described above, wild boars are rather territorial, allowing the geographical allocation of such data. Only the renegade wild sows are known to travel across large areas when they are searching for a new herd. A study from the Czech Republic, traditionally a country with a high TBE incidence, found a positive association between the number of hunted wild boar and human cases. Consequently, the authors concluded that wild boar must play a role in TBEV transmission either directly or indirectly.89
In Finland, moose (Alces alces) and white-tailed deer (Odocoileus virginianus) were found to harbor TBEV-specific antibodies (0.74%) and the use of such seroprevalence data as an indicator for local risk of human TBE infection is recommended.90 In Norway, 9.4% of 286 moose, 1.4% in red deer and 0.7% in roe deer led to an overall seroprevalence of 4.6% in cervids. Interestingly none of the 83 investigated reindeer showed antibodies against TBEV.91 One single case report describes the pathological and immunohistological findings in a mouflon (Ovis ammon musimon) with marked encephalitis due to TBEV.92 A Polish study analyzed D. reticulatus collected from the lowland European bison (Bison bonasus bonasus) in a known endemic focus and found 18.42% of these ticks to be positive for TBEV RNA.93 Seroprevalence in the bison themselves was found to be >60%.94 In Japan, the seroprevalence in raccoons varied between 0.8% and 5.9% in eastern and central Hokkaido province while sika deer (Cervus Nippon) showed in TBEV-neutralizing antibodies in 0.8% and 2.4% there.95 A recent case report of a sickened chamois in Italy is further challenging our view that wild animals do not suffer upon infection with TBEV and only seroconvert.96 It might well be that we have not looked close enough in the past in sick and deceased wildlife. Interestingly, not much is known about the role of foxes (Vulpes vulpes) in natural TBE foci. Although it is a highly prevalent predator of small mammals (see below), and is regularly infested with Ixodes ticks, there are no recent studies investigating virus or antibodies against TBEV in foxes. Older studies from Germany were mostly performed in non-endemic areas on the German-Dutch border and Brandenburg, and consequently revealed no seroprevalence or a single sero-reactive serum sample only.97,98 However, the latter report found every third fox in South-Western Germany to have antibodies against TBEV.98 In another study in Germany with a large sample size of more than 1200 fox sera, an overall prevalence >20% was found, again correlating with TBE endemic areas (>30% positive samples) versus non-endemic areas with just 13% seropositive fox samples.99 In Croatia, a study found at least 1.6% of ticks on red foxes and 1.1% of spleen samples of red deer (Cervus elaphus) to be positive for TBEV-RNA.100 It would be interesting and necessary to perform a seroprevalence study in a known endemic area to shed light on the role of the fox in the natural transmission cycle of TBEV and to prove the putative positive correlation between fox abundance and TBE incidence.98,101
Studies trying to detect a correlation between human TBE incidence and the abundance of certain animals are contradictory. A Swedish study revealed that, with one year of time-lag, the abundance of roe deer, red deer, mountain hare (Lepus timidus) and European hare (Lepus europaeus) showed positive covariance with the incidence of human TBE.102 In contrast, moose and fallow deer (Dama dama) showed negative covariance and wild boar, lynx (Lynx lynx) and red fox showed no significant covariance with human TBE incidence.99 In Slovenia, red deer abundance was correlated with human TBE incidence when including a three-year time-lag, whereas roe deer showed no significant correlation.103 An Italian study found roe deer abundance to have a better predictive value for a model explaining the increasing human TBE incidence than roe deer abundance.104
Small mammals
Small mammals have an essential role in the maintenance of TBE foci in 2 ways. Firstly, rodents and shrews are the main hosts for Ixodes larvae. Without this first blood meal, a tick population would die out over time. They are also, to a lesser extent, hosts for nymphs when they take their blood meal, which is needed before they can molt into adult ticks. Secondly, they are reservoir hosts for TBEV and thus responsible for infections of ticks. The reservoir function, however, has large implications for the longevity of a natural focus. As outlined earlier, in the chapter on transmission and natural cycle, infection of a tick can occur via a viremic host, but another phenomenon has been described which also applies to the infection of ticks while feeding on small mammals. The so-called co-feeding allows the infection of Ixodes larvae when an infected Ixodes nymph feeds in close proximity. In this case, the rodent does not have to be infected, because the virus finds its way from the nymph directly to the larva.105 So, it is safe to say that, in many ways, rodents are as necessary as Ixodes ticks for maintaining the TBEV life-cycle. In particular bank voles (Clethrionomys glareolus) appear to be well adapted to TBEV, leading to long-lasting viremias and infiltration of the brain without causing visible neurological symptoms.106
Recent publications have reviewed the prevalence of either viral ribonucleic acid (RNA) or specific antibodies against TBEV in rodents in various countries.107-110 The antibody prevalence in endemic areas was found to range between 0% and 5.9%. However, seroprevalence rates up to 12.5% were found in some rodent species (e.g., the bank vole, Clethrionomys glareolus),111 suggesting a differing role of particular rodent species in a TBE focus. Viral RNA can also be found in wild rodents, with an even higher prevalence of up to 15%.112 Studies from Hungary identified TBEV-RNA in 4.2%113 and TBEV-specific anti-bodies in 5.2% and 4.9% of the tested small rodents.114 Recently, TBEV-positive bank voles (and ticks) were found in a forest within the city borders of Moscow, Russia.115 Experimentally infected common voles (Microtus arvalis) harbored infectious TBEV for at least 3 months.112 Viral RNA could be found in the brain tissue of experimentally infected bank voles for up to 168 days.116 This has important implications, as the brain (and to a lesser extent other organs such as kidney and spleen) seems to be the prime site of virus persistence in rodents. Indeed, TBE viral RNA was found in the brain tissue of naturally infected field voles (Microtus agrestis) and bank voles in Finland, after the winter but before the tick season started.117 Seroprevalence in Microtus rodents were found to be 4% in Poland.118 Thus rodents seem, along with transstadially-infected ticks, to play a role in the ‘overwintering’ of the TBEV.
Other mammals and birds
As most animals do not develop overt disease upon infection with TBEV, many mammal species have never been investigated as to whether or not they are susceptible to an infection or capable of developing an immune response in terms of measurable antibody titers. According to the broad geographic distribution of TBE covering most of Europe and northern Asia, we consider that there may be many mammal species not yet investigated that react to an infection in a similar manner as described above for wild boars or roe deer, i.e., seroconversion without clinical disease. One exception is the Barbary macaque (Macaca sylvanus), a monkey species not native to Eurasia, despite a small population in Gibraltar, the southernmost tip of Spain. An individual of a small group of these animals kept in southwest Germany in an outdoor area fell severely ill with central nervous symptoms and was euthanized for ethical reasons. A pan-encephalitis was diagnosed and TBEV was demonstrated by immunohistochemistry, real-time RT-PCR, and virus isolation.119,120 Other individuals of this monkey group sero-converted without showing clinical signs.121 Thus far, we are not aware of further case reports of non-native species kept in semi-free holdings or zoos.
Birds are known to be readily infested with ixodid ticks and are prime suspects for long-distance transportation of ticks.122 The first studies investigating the prevalence of TBEV-harboring ticks on birds came from the Ottenby Bird Observatory at the southern tip of the island Öland in Sweden. During the annual ringing, more than 1000 Ixodes spp. ticks were collected from birds, with 0.52% showing TBEV RNA.123 Subsequent studies from Estonia (0.4% positive nymphs),124 Switzerland (0.27% TBE viral RNA positive),125 Latvia (14%),126 Germany (no TBE virus found in almost 2500 Ixodes ricinus ticks collected from birds)122 and Slovakia127 (a brain sample in a buzzard, Buteo buteo) demonstrated the possibility that TBEV can be transported over rather long distances via infected ticks attached to birds.
Studies from the 1960s failed to demonstrate both viremia and clinical illness in great tits (Parus major), pheasants (Phasianus colchicus), falcons (Falco tinnunculus), and buzzards (Buteo buteo).51 Only a small fraction of infected animals seroconverted. Other birds, such as the house sparrow (Passer domesticus), common redpoll (Acanthis flammea), quail (Coturnix coturnix), and duck (Anas platyrhynchos), showed either detectable virus or even moderate viremia after infection.128 Another study demonstrated that the presence of TBEV seems to vary according to season and bird species. Prevalence rates above 50% indicate that particular bird species like fieldfares (Turdus pilaris), bramblings (Fringilla montifrigilla), and the common redstart (Phoenicurus phoenicurus) may well play a role as a reservoir, or at least amplifying host, for TBEV.129
Veterinary diagnostic aspects
In general, the same diagnostic tests and methods are applied for animals as those that are currently in use for diagnostic purposes in humans (see Chapter 11: Diagnostic of TBEV-infections). With the exception of diseased dogs and horses, which are usually under tight supervision by their owner, the time window to use any direct detection method for TBEV – isolation or real-time RT-PCR – is usually too short to be of any practical relevance. Immuno-histochemistry may be used in euthanized animals. In epidemiological studies using rodents, these methods may be applied as virus and viral RNA can be detected in the brain tissue of infected animals for months (see above). In contrast, serology can be easily applied in any animal species. Three test formats are frequently used for this purpose, i.e., ELISA, IFA (immunofluorescence assay), and SNT. The ELISA can be performed with a species-specific conjugate, which is available for dogs, cattle, sheep, goats, swine (works also for wild boar), cervids, and mouse (works also for voles and mice). However, there is a commercially available, species-independent ELISA which uses protein G-coupled enzyme. Although this test is also available for immunoglobulin (Ig)M antibodies, the IgG version should be used because of the reasons mentioned above. The IFA usually uses a mixture of uninfected and TBEV-infected Vero cells fixed on slides and the antibody-conjugates described for the ELISA. Finally, the SNT is the gold standard and is needed in order to verify results of the other 2 assays. According to the European Centre for Disease Prevention and Control, an SNT titer =1:10 confirms the diagnosis.35,39,130
Concluding remarks
Infections of various animals with TBEV are common in TBE-endemic areas, although they are barely noticed due to the lack of overt disease. The known exceptions are dogs and horses, which can become severely ill with the same panel of clinical symptoms, as the same neurological regions in the CNS are affected. Domestic ruminants are a risk for human health as they can shed TBEV through their milk for many days. If unpasteurized, TBEV-contaminated milk or milk products are ingested by consumers, clusters of human cases may be the consequence. Many wild animal species become infected and develop an antibody response, but they do not appear to be harmed. However, some recently published cases of clinical TBE in a single roe dear and a single chamois challenge this view. Future research may address the potential use of antibody prevalence rates of particular animal species in order to complement the current risk definition for human infections, which at the moment is largely based on the count of human cases alone. Finally, birds seem to play a role in long-distance transportation of TBEV-infected ticks and thus the geographic spread, while small mammals, in particular rodents, are the key players in maintaining a TBE focus in nature.
Contact
Martin Pfeffer
pfeffer@vetmed.uni-leipzig.de
Authors
Martin Pfeffer, Hannah M. Schmuck, Michael Leschnik
Citation
Pfeffer M, Schmuck HM, Leschnik M. TBE in animals. Chapter 10. In: Dobler G, Erber W, Bröker M, Chitimia-Dobler L, Schmitt HJ, eds. The TBE Book. 7th ed. Singapore: Global Health Press; 2024. doi: 10.33442/26613980_10-7
References
- Lindquist L, Vapalahti O. Tick-borne encephalitis. Lancet. 2008;371:1861-71.
- Dobler G, Gniel D, Petermann R, Pfeffer M. Epidemiology and distribution of tick-borne encephalitis. Wien Med Wochenschr. 2012;162:230-8.
- Leschnik M, Feiler A, Duscher G, Joachim A. Effect of owner- controlled acaricidal treatment on tick infestation and immune response to tick-borne pathogens in naturally infested dogs from Eastern Austria. Parasit Vectors. 2013;6:62.
- Matile H, Ferrari E, Aeschlimann, A, Wyler R. Die Verbreitung der Zecken-enzephalitis in der Schweiz. Schweiz Med Wochenschr. 1981;111:1262-9.
- Chambouris R, Sixl W, Stunzer D, Köck M. Antibodies in dogs to the virus of tick-borne encephalitis (early summer encephalomyelitis/tick-borne encephalitis) in Greece. Geogr Med. 1989;3:11-14.
- Balling A, Beer M, Gniel D, Pfeffer M. Prevalence of antibodies against tick-borne encephalitis virus in dogs from Saxony, Germany. Berl Münch Tierärztl Wochenschr. 2015;128:297- 303.
- Reiner B, Grasmück S, Steffen F, et al. Prevalence of TBE antibodies in serum and CSF of dogs with inflammatory and non-inflammatory CNS disease. Int J Med Microbiol. 2002;291 (Suppl. 33):234.
- Roelandt S, Heyman P, Filette MD, et al. Tick-Borne Encephalitis Virus Seropositive Dog Detected in Belgium: Screening of the Canine Population as Sentinels for Public Health. Vector Borne Zoonotic Dis. 2011;11:1371-6.
- Lindhe KES, Meldgaard DS, Jensen PM, Houser GA, Berendt M. Prevalence of tick-borne encephalitis virus antibodies in dogs from Denmark. Acta Vet Scand. 2009;51:56.
- Klimeš J, Juřicová Z, Literák I, Schánilec P, Trachta e Silva E. Prevalence of antibodies to tick-borne encephalitis and West Nile flaviviruses and the clinical signs of tick-borne encephalitis in dogs in the Czech Republic. Vet Rec. 2001;148:17-20.
- Hekrlová A, Kubíček O, Lány P, Rosenbergová K, Schánilec P. Tick-borne encephalitis in dogs: application of ‘nested real- time RT-PCR’ for intraviral virus detection. Berl Münch Tierärztl Wochenschr. 2015;128:397-401.
- Csángó PA, Blakstad E, Kirtz GC, Pedersen JE, Czettel B. Tick- borne Encephalitis in Southern Norway. Emerg Infect Dis. 2004;10:533-4.
- Levanov L, Perez Vera C, Vapalahti O. Prevalence estimation of tick-borne encephalitis virus (TBEV) antibodies in dogs from Finland using novel dog anti-TBVE IgG Mab-capture and IgG immunofluorescence assays based on recombinant TBEV subviral particles. Ticks Tick-Borne Dis. 2016;7:979-82.
- Kirtz G, Kölbl S, Czettel B, Thalhammer JG Frühsommer- Meningoenzephalitis (FSME, Zetraleuropäische Zeckenenzephalitis) beim Hund in Österreich: eine Seroprävalenz-Studie. Kleintierpraxis. 2003;48:133-40.
- Wójcik-Fatla A, Cisak E, Zajac V, Zwoliński J, Dutkiewicz J. Prevalence of tick-borne encephalitis virus in Ixodes ricinus and Dermacentor reticulatus ticks collected from the Lublin region (eastern Poland). Ticks Tick-Borne Dis. 2011;2: 16-19.
- Nygren TM, Pilic A, Böhmer MM, Wagner-Wiening C, Wichmann O, Harder T, Hellenbrand W Tick-Borne Encephalitis Risk Increases with Dog Ownership, Frequent Walks, and Gardening: A Case-Control Study in Germany 2018-2020. Microorganisms. 2022 Mar 23;10(4):690. doi: 10.3390/microorganisms10040690.
- Girl P, Haut M, Riederer S, Pfeffer M, Dobler G. Comparison of Three Serological Methods for the Epidemiological Investigation of TBE in Dogs. Microorganisms. 2021 Feb 15;9(2):399. doi: 10.3390/microorganisms9020399.
- Pfeffer M, Dobler G. Tick-borne encephalitis virus in dogs–is this an issue? Parasit Vectors. 2011 Apr 13;4:59. doi: 10.1186/1756-3305-4-59.
- Kleeb C, Golini L, Beckmann K, Torgerson P, Steffen F. Canine Tick-Borne Encephalitis: Clinical Features, Survival Rate and Neurological Sequelae: A Retrospective Study of 54 Cases (1999-2016). Front Vet Sci. 2021 Nov 10;8:782044. doi: 10.3389/fvets.2021.782044. eCollection 2021.
- Potkonjak A, Petrović T, Ristanović E, Lalić I, Vračar V, Savić S, Turkulov V, Čanak G, Milošević V, Vidanović D, Jurišić A, Petrović A, Petrović V. Molecular Detection and Serological Evidence of Tick-Borne Encephalitis Virus in Serbia. Vector Borne Zoonotic Dis. 2017 Dec;17(12):813-820. doi: 10.1089/vbz.2017.2167.
- Salat J, Hunady M, Schanilec P, Strakova P, Stefanik M, Svoboda P, Strelcova L, Bojcukova J, Palus M, Růžek D. Experimental and Natural Infections of Tick-Borne Encephalitis Virus in Dogs. Viruses. 2021 Oct 9;13(10):2039. doi: 10.3390/v13102039.
- Topp AK, Springer A, Mischke R, Rieder J, Feige K, Ganter M, Nagel-Kohl U, Nordhoff M, Boelke M, Becker S, Pachnicke S, Schunack B, Dobler G, Strube C. Seroprevalence of tick-borne encephalitis virus in wild and domestic animals in northern Germany. Ticks Tick Borne Dis. 2023 Nov;14(6):102220. doi: 10.1016/j.ttbdis.2023.102220.
- Simkute E, Pautienius A, Grigas J, Urbute P, Stankevicius A. The Prevalence, Seroprevalence, and Risk Factors of Tick-Borne Encephalitis Virus in Dogs in Lithuania, a Highly Endemic State. Viruses. 2023 Nov 17;15(11):2265. doi: 10.3390/v15112265.
- Andersson E, Kendall A, Url A, Auer A, Leschnik M. The first RT-qPCR confirmed case of tick-borne encephalitis in a dog in Scandinavia. Acta Vet Scand. 2020 Sep 10;62(1):51. doi: 10.1186/s13028-020-00550-2.
- Dultz R, Goldhammer M. Tick-borne encephalitis in a dog. Tierarztl Prax Ausg K Kleintiere Heimtiere. 2021 Oct;49(5):377-381. doi: 10.1055/a-1580-8386. [in German]
- Sioutas G, Tsakou K, Top C, Jongejan F, Papadopoulos E. First clinical case of tick-borne encephalitis (TBE) in a dog in Greece. Ticks Tick Borne Dis. 2023 Nov;14(6):102226. doi: 10.1016/j.ttbdis.2023.102226.
- Leschnik MW, Kirtz GC, Thalhammer JG. Tick-borne encephalitis (TBE) in dogs. Int J Med Microbiol. 2002;291 (Suppl. 33):66-9.
- Leschnik MW, Benetka V, Url A, et al. Virale Enzephalitiden beim Hund in Österreich: diagnostische und epidemiologische Aspekte. Vet Med Austria / Wien Tierärztl Monatsschr. 2008;95:190-9.
- Reiner B, Fischer A, Gödde T, Müller W. Clinical diagnosis of canine tick-borne encephalitis (tbe): Contribution of cerebrospinal fluid analysis (csf) and csf antibody titers. Zent Bl Bakteriol. 1999;289:605-9.
- Leschnik M. Tick-borne encephalitis in dogs. Proc. Abildgaard Symposium, Kopenhagen, Dänemark. 2005:43-5.
- Tipold A, Fatzer R, Holzmann H. Zentraleuropäische Zeckenencephalitis beim Hund. Kleintierpraxis. 1993;38:619- 28.
- Stadtbäumer K, Leschnik MW, Nell B. Tick-borne encephalitis virus as a possible etiologic agent for optic neuritis in a dog. Vet Ophthal. 2004;7:271-7.
- Levanov L, Vera CP, Vapalahti O. Prevalence estimation of tick-borne encephalitis virus (TBEV) antibodies in dogs from Finland using novel dog anti-TBEV IgG MAb-capture and IgG immunofluorescence assays based on recombinant TBEV subviral particles. Ticks Tick Borne Dis. 2016 Jul;7(5):979-982. doi: 10.1016/j.ttbdis.2016.05.002.
- Alnefelt Y, Van Meervenne S, Varjonen K, Tidholm A, Rohdin C. Evaluation of antibodies in cerebrospinal fluid for the diagnosis of tick-borne encephalitis in dogs. Acta Vet Scand. 2021 Aug 26;63(1):32. doi: 10.1186/s13028-021-00597-9.
- Klaus C, Ziegler U, Kalthoff D, Hoffmann B, Beer M. Tick-borne encephalitis virus (TBEV) – findings on cross reactivity and longevity of TBEV antibodies in animal sera. BMC Vet Res. 2014;10:78.
- Beckmann K, Steffen F, Ohlerth S, Kircher PR, Carrera I. Three tesla magnetic resonance imaging findings in 12 cases of canine central European tick-borne meningoencephalomyelitis. Vet Radiol Ultrasound. 2016;57:41-8.
- Weissenböck H, Suchy A, Holzmann H. Tick-borne encephalitis in dogs: neuropathological findings and distribution of antigen. Acta Neuropathol. 1998;95: 361-6.
- Sievert C, Hening R, Beckmann K, Kircher PR, Carrera I. Comparison between proton magnetic resonance spectroscopy findings in dogs with tick-borne encephalitis and clinically normal dogs. Vet Radiol Ultrasound. 2016;58:53-61.
- Klaus C, Beer M, Saier R, Schubert H, Bischoff S, Süss J. Evaluation of serological tests for detecting tick-borne encephalitis virus (TBEV) antibodies in animals. Berl Münch Tierärztl Wochenschr. 2011;124:443-9.
- Waldvogel K, Matile H, Wegmann C, Wyler R, Kunz C. Zeckenenzephalitis beim Pferd. Schweiz Arch Tierheilkd. 1981;123:227-33.
- Luckschander N, Kölbl S, Enzensberger O, Zipko HT, Thalhammer JG. Tick-borne encephalitis (TBE) in an Austrian horse population. Tierärztl Prax Ausgabe G. 1999;27:235-8.
- Müller K, König M, Thiel HJ. [Tick-borne encephalitis (TBE) with special emphasis on infection in horses]. Dtsch Tierärztl Wochenschr. 2006;113:147-51 [in German].
- Klaus C, Hörügel U, Beer M. Tick-borne encephalitis virus (TBEV) infection in horses: Clinical and laboratory findings and epidemiological investigations. Vet Microbiol. 2013;163:368- 72.
- de Heus P, Bagó Z, Weidinger P, Lale D, Trachsel DS, Revilla-Fernández S, Matiasek K, Nowotny N. Severe Neurologic Disease in a Horse Caused by Tick-Borne Encephalitis Virus, Austria, 2021. Viruses. 2023 Sep 29;15(10):2022. doi: 10.3390/v15102022.
- Rushton JO, Lecollinet S, Hubálek Z, et al. Tick-borne encephalitis virus in horses, Austria, 2011. Emerg Infect Dis. 2013;19:635-7.
- Ganzenberg S, Sieg M, Ziegler U, Pfeffer M, Vahlenkamp TW, Hörügel U, Groschup MH, Lohmann KL. Seroprevalence and Risk Factors for Equine West Nile Virus Infections in Eastern Germany, 2020. Viruses. 2022 May 30;14(6):1191. doi: 10.3390/v14061191.
- Gothe LMR, Ganzenberg S, Ziegler U, Obiegala A, Lohmann KL, Sieg M, Vahlenkamp TW, Groschup MH, Hörügel U, Pfeffer M. Horses as Sentinels for the Circulation of Flaviviruses in Eastern-Central Germany. Viruses. 2023 Apr 30;15(5):1108. doi: 10.3390/v15051108.
- Janitza-Futterer D. Serologische Untersuchungen zur endemischen Situation der Infektion mit dem FSME-Virus in einer südbadischen Pferde- und Hundepopulation. Vetmed Diss Munich. 2003.
- Sikutova S, Hornok S, Hubalek Z, et al. Serological survey of domestic animals for tick-borne encephalitis and Bhanja viruses in northeastern Hungary. Vet Microbiol. 2009;135:267 -71.
- Sedlák K, Zelená H, Křivda V, Šatrán P. Surveillance of West Nile fever in horses in the Czech Republic from 2011 to 2013. Epidemiol Mikrobiol Immunol. 2014; 63:307-11.
- Camino E, Schmid S, Weber F, Pozo P, de Juan L, König M, Cruz-Lopez F. Detection of antibodies against tick-borne encephalitis flaviviruses in breeding and sport horses from Spain. Ticks Tick Borne Dis. 2020 Sep;11(5):101487. doi: 10.1016/j.ttbdis.2020.101487.
- Vilibic-Cavlek T, Krcmar S, Bogdanic M, Tomljenovic M, Barbic L, Roncevic D, Sabadi D, Vucelja M, Santini M, Hunjak B, Stevanovic V, Boljfetic M, Bjedov L, Masovic V, Potocnik-Hunjadi T, Lakoseljac D, Al-Mufleh M, Savic V. An Overview of Tick-Borne Encephalitis Epidemiology in Endemic Regions of Continental Croatia, 2017-2023. Microorganisms. 2024 Feb 13;12(2):386. doi: 10.3390/microorganisms12020386.
- Pautienius A, Armonaite A, Simkute E, Zagrabskaite R, Buitkuviene J, Alpizar-Jara R, Grigas J, Zakiene I, Zienius D, Salomskas A, Stankevicius A. Cross-Sectional Study on the Prevalence and Factors Influencing Occurrence of Tick-Borne Encephalitis in Horses in Lithuania. Pathogens. 2021 Jan 31;10(2):140. doi: 10.3390/pathogens10020140.
- Cleton NB, van Maanen K, Bergervoet SA, et al. A serological protein microarray for detection of multiple cross-reactive flavivirus infections in horses for veterinary and public health surveillance. Transbound Emerg Dis. 2017; 64:1801–12
- Gresiková M. Excretion of the tick-borne encephalitis virus in the milk of subcutaneously infected cows. Acta Virol. 1958;2:188-92.
- Nosek J, Kozuch O, Ernek E, Lichard M. The importance of goats in the maintenance of tick-borne encephalitis virus in nature. Acta Virol. 1967;11:470-82.
- Böhm B, Schade B, Bauer B, Hoffmann B, Hoffmann D, Ziegler U, Beer M, Klaus C, Weissenböck H, Böttcher J. Tick-borne encephalitis in a naturally infected sheep. BMC Vet Res. 2017;13:267.
- Balogh Z, Egyed I, Ferenczi E, et al. Experimental infection of goats with tick-borne encephalitis virus and possibilities to prevent virus transmission by raw goat milk. Intervirology. 2012;55:194-200.
- Rieille N, Klaus C, Hoffmann D, Péter O, Voordouw M. Goats as sentinel hosts for the detection of tick-borne encephalitis risk areas in the Canton of Valais, Switzerland, BMC Vet Res. 2017;13:217.
- Casati Pagani S, Frigerio Alossa S, Klasu C, Hoffmann D, Beretta O, Bomio-Pacciorini N, Lazzaro M, Merlani G, Ackermann R, Beuret C. First detection of TBE virus in ticks and sero-reactivity in goats in a non-endemic region in the southern part of Switzerland (Canton of Ticino). Ticks Tick-Borne Dis. 2019;10:868-874.
- Bauer BU, Könenkamp L, Stöter M, Wolf A, Ganter M, Steffen I, Runge M. Increasing awareness for tick-borne encephalitis virus using small ruminants as suitable sentinels: Preliminary observations. One Health. 2021 Feb 20;12:100227. doi: 10.1016/j.onehlt.2021.100227.
- Wallenhammar A, Lindqvist R, Asghar N, Gunaltay S, Fredlund H, Davidsson Å, Andersson S, Överby AK, Johansson M. Revealing new tick-borne encephalitis virus foci by screening antibodies in sheep milk. Parasit Vectors. 2020 Apr 8;13(1):185. doi: 10.1186/s13071-020-04030-4.
- Dorko E, Hockicko J, Rimárová K, Bušová A, Popad’ák P, Popad’áková J, Schréter J. Milk outbreaks of tick-borne encephalitis in Slovakia, 2012-2016. Cent Eur J Public Health 2018;26(Suppl):S47-S50.
- Kerlik J, Avdičová M, Štefkovičová M, Tarkovská V, Pántiková Valachová M, Molčányi T, Mezencev R. Slovakia reports highest occurrence of alimentary tick-borne encephalitis in Europe: Analysis of tick-borne encephalitis outbreaks in Slovakia during 2007-2016. Travel Med Infect Dis. 2018;26:37-42.
- Markovinović L, Kosanović Ličina ML, Tešić V, Vojvodić D, Vladušić Lucić I, Kniewald T, Vukas T, Kutleša M, Krajinović LC. An outbreak of tick-borne encephalitis associated with raw goat milk and cheese consumption, Croatia, 2015. Infection. 2016 Oct;44(5):661-5. doi: 10.1007/s15010-016-0917-8.
- Ilic M, Barbic L, Bogdanic M, Tabain I, Savic V, Kosanovic Licina ML, Kaic B, Jungic A, Vucelja M, Angelov V, Kovacevic M, Roncevic D, Knezevic S, Stevanovic V, Slavuljica I, Lakoseljac D, Vickovic N, Bubonja-Sonje M, Hansen L, Vilibic-Cavlek T. Tick-borne encephalitis outbreak following raw goat milk consumption in a new micro-location, Croatia, June 2019. Ticks Tick Borne Dis. 2020 Nov;11(6):101513. doi: 10.1016/j.ttbdis.2020.101513.
- Brockmann SO, Oehme R, Buckenmaier T, Beer M, Jeffery-Smith A, Spannenkrebs M, Haag-Milz S, Wagner-Wiening C, Schlegel C, Fritz J, Zange S, Bestehorn M, Lindau A, Hoffmann D, Tiberi S, Mackenstedt U, Dobler G.
A cluster of two human cases of tick-borne encephalitis (TBE) transmitted by unpasteurised goat milk and cheese in Germany, May 2016. Euro Surveill. 2018;23(15):pii=17-00336. - Cisak E, Wójcik-Fatla A, Zajac V, et al. Prevalence of tick-borne encephalitis virus (TBEV) in samples of raw milk taken randomly from cows, goats and sheep in eastern Poland. Ann Agric Env Med. 2010;17:283-6.
- Paulsen KM, Stuen S, das Neves CG, Suhel F, Gurung D, Soleng A, Stiasny K, Vikse R, Andreassen ÅK, Granquist EG. Tick-borne encephalitis virus in cows and unpasteurized cow milk from Norway. Zoonoses Public Health. 2019;66:212-222.
- Klaus C, Ziegler U, Hoffmann D, Press F, Fast C, Beer M. Tick-borne encephalitis virus (TBEV) antibodies in animal sera – occurence in goat flocks in Germany, longevits and ability to recall immunological information after more than six years. BMC Veterinary Res. 2019;15:399.
- Klaus C, Hoffmann B, Moog U, et al. Can goats be used as sentinels for Tick-borne encephalitis (TBE) in non-endemic areas? Experimental studies and epizootiological observations. Berl Münch Tierärztl Wochenschr. 2010;123:441-5.
- Klaus C, Beer M, Saier R, et al. Goats and sheep as sentinels for tick-borne encephalitis (TBE) virus – Epidemiological studies in areas endemic and non-endemic for TBE virus in Germany. Ticks Tick-Borne Dis. 2012;3:27-37.
- Klaus C, Hoffmann D, Hoffmann B, Beer M. Frühsommer- Meningoenzephalitis-Virus-Infektionen bei Tieren – Klinik, Diagnostik und epidemiologische Bedeutung. Berl Münch Tierärztl Wochenschr. 2016;130:102-12.
- Vor T, Kiffner C, Hagedorn P, Niedrig M, Ruhe F. Tick burden on European roe deer (Capreolus capreolus). Exp Appl Acarol. 2010;51:405-17.
- Jahfari S, de Vries A, Rijks JM, van Gucht S, Vennema H, Sprong H, Rockx B. Tick-Borne Encephalitis Virus in Ticks and Roe Deer, the Netherlands, Emerg Infect Dis. 2017;23:1028–1030.
- Da Rold G, Obber F, Monne I, Milani A, Ravagnan S, Toniolo F, Sgubin S, Zamperin G, Foiani G, Vascellari M, Drzewniokova P, Castellan M, De Benedictis P, Citterio CV. Clinical Tick-Borne Encephalitis in a Roe Deer (Capreolus capreolus L.). Viruses. 2022 Jan 31;14(2):300. doi: 10.3390/v14020300.
- Gerth HJ, Grimshandl D, Stage B, Döller G, Kunz C. Roe deer as sentinels for endemicity of tick-borne encephalitis virus. Epidemiol Infect. 1995;115:355-65.
- Van der Poel WH, Van der Heide R, Bakker D, et al. Attempt to detect evidence for tick-borne encephalitis virus in ticks and mammalian wildlife in the Netherlands. Vector Borne Zoonotic Dis. 2005;5:58-64.
- Skarphédinsson S, Jensen PM, Kristiansen K. Survey of tick-borne infections in Denmark. Emerg Infect Dis. 2005;11:1055-61.
- Kiffner C, Vor T, Hagedorn P, Niedrig M, Rühe F. Determinants of tick-borne encephalitis virus antibody presence in roe deer (Capreolus capreolus) sera. Med Vet Entomol. 2012;26:18-25.
- Ytrehus B, Vainio K, Dudman SG, Gilray J. Tick-borne encephalitis virus and Louping-Ill virus may co-circulate in southern Norway. Vector Borne Zoonotic Dis. 2013;13:762-8.
- Balling A, Plessow U, Beer M, Pfeffer M. Prevalence of antibodies against tick-borne encephalitis virus in wild game from Saxony, Germany. Ticks Tick Borne Dis. 2014;5:805-9.
- Duscher GG, Wetscher M, Baumgartner R. Roe deer sera used for TBE surveillance in Austria. Ticks Tick Borne Dis. 2015;6:489-93.
- Frimmel S, Leister M, Löbermann M, Feldhusen F, Seelmann M, Süss J, Reisinger EC. Seroprevalence of tick-borne-encephalitis virus in wild game in Mecklenburg-Western Pomerania (north-eastern Germany). Ticks Tick Borne Dis. 2016;7:1151–4.
- Roelandt S, Suin V, Van der Stede Y, et al. First TBEV serological screening in Flemish wild boar. Infect Ecol Epidemiol. 2016;6:31099.
- Donoso Mantke O, Escadafal C, Niedrig M, Pfeffer M. Tick-borne encephalitis in Europe, 2007 to 2009. Euro Surveill. 2011;16:pii=19976.
- Kunze U, the ISW-TBE. Tick-borne encephalitis – still on the map. Report of the 18th annual meeting of the international scientific working group on tick-borne encephalitis (ISW-TBE). Ticks Tick Borne Dis. 2016;7:911-4.
- Bournez L, Umhang G, Faure E, Boucher J-M, Boué F, Jourdain E, Sarasa M, Llorente F, Jiménez-Clavero MA, Moutailler S, Lacour SA, Lecollinet S, Beck C. Exposure of wild ungulates to the Usutu and Tick-borne encephalitis viruses in France in 2009-2014: Evidence of undetected flavivirus circulation a decade ago. viruses 2020;12:10.
- Kríz B, Daniel M, Benes C. The role of game (wild boar and roe deer) in the spread of tick-borne encephalitis in the Czech Republic. Vector Borne Zoonotic Dis. 2014;14:801-7.
- Tonteri E, Jokelainen P, Matala J, Pusenius J, Vapalahti O. Serological evidence of tick-borne encephalitis virus infection in moose and deer in Finland: sentinels for virus circulation. Parasit Vectors. 2016;9:54.
- Paulsen KM, das Neves CG, Granquist EG, Madslien K, Stuen S, Pedersen BN, Viske R, Rocchi M, Laming E, Stiasny K, Andreassen ÅK. Cervids as sentinel-species for tick-borne encephalitis virus in Norway – a serological study. Zoonoses Public Health. 2019 Dec 19;[online ahead of print]
- Bagó Z, Bauder B, Kolodziejek J, Nowotny N, Weissenböck H. Tick-borne encephalitis in a mouflon (Ovis ammon musimon). Vet Rec. 2002;150:218-20.
- Biernat B, Karbowiak G, Stańczak J, Masny A, Werszko J. The first detection of the tick-borne encephalitis virus (TBEV) RNA in Dermacentor reticulatus ticks collected from the lowland European bison (Bison bonasus L.). Acta Parasitologica. 2016;61:130–5.
- Krzysiak MK, Anusz K, Konieczny A, Rola J, Salat J, Strakova P, Olech W, Larska M. The European bison (Bison bonasus) as an indicatory species for the circulation of tick-borne encephalitis virus (TBEV) in natural foci in Poland. Ticks Tick Borne Dis. 2021 Nov;12(6):101799. doi: 10.1016/j.ttbdis.2021.101799.
- Jamsransuren D, Kentaro Y, Hiroaki K, Mitsuhiko A, Kei O, Kei F, et al. Epidemiological survey of tick-borne encephalitis virus infection in wild animals an Hokkaido and Honshu islands, Japan. Japanese J Veterinary Res. 2019;67:163-172.
- Gaffuri A, Sassera D, Calzolari M, Gibelli L, Lelli D, Tebaldi A, Vicari N, Bianchi A, Pigoli C, Cerioli M, Zandonà L, Varisco G, Bertoletti I, Prati P. Tick-Borne Encephalitis, Lombardy, Italy. Emerg Infect Dis. 2024 Feb;30(2):341-344. doi: 10.3201/eid3002.231016.
- Wurm R, Dobler G, Peters M, Kiessig ST. Serological investigations of red foxes (Vulpes vulpes L.) for determination of the spread of tick-borne encephalitis in North Rhine-Westphalia. J Vet Med B Infect Vet Public Health. 2000;47:503-9.
- Rieger MA, Nübling M, Müller W, Hasselhorn HM, Hofmann F. Foxes as indicators for TBE endemicity – a comparative serological and investigation. Zentralbl Bakteriol. 1999;289:610-18.
- Haut M, Girl P, Oswald B, Romig T, Obiegala A, Dobler G, Pfeffer M. The Red Fox (Vulpes vulpes) as Sentinel for Tick-Borne Encephalitis Virus in Endemic and Non-Endemic Areas. Microorganisms. 2020 Nov 18;8(11):1817. doi: 10.3390/microorganisms8111817.
- Jemeršić L, Dežđek D, Brnić D, Prpić J, Janicki Z, Keros T, Roić B, Slavica A, Terzić S, Konjević D, Beck R. Detection and genetic characterization of tick-borne encephalitis virus (TBEV) derived from ticks removed from red foxes (Vulpes vulpes) and isolated from spleen samples of red deer (Cervus elaphus) in Croatia. Ticks Tick Borne Dis. 2014;5:7-13.
- Haemig PD, Lithner S, Sjöstedt de Luna, S, et al. Red fox and tick-borne encephalitis (TBE) in humans: Can predators influence public health. Scand J Infect Dis. 2008;40:527-32.
- Jaenson TGT, Petersson EH, Jaenson DGE, Kindberg J, Pettersson JH, Hjertqvist M, Medlock JM, Bengtsson H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: incidence of human TBE correlates with abundance of deer and hares, Parasit Vectors. 2018;11:477.
- Knap N, Avšič-Županc T. Correlation of TBE Incidence with Red Deer and Roe Deer Abundance in Slovenia. PLoS ONE. 2013;8:e66380.
- Rizzoli A, Hauffe HC, Tagliapietra V, Neteler M, Rosà R Forest Structure and Roe Deer Abundance Predict Tick-Borne Encephalitis Risk in Italy. PLoS ONE. 2009;4:e4336.
- Labuda M, Kozuch O, Zuffová E, et al. Tick-borne encephalitis virus transmission between ticks cofeeding on specific immune natural rodent hosts. Virology. 1997; 235:138-43.
- Michelitsch A, Tews BA, Klaus C, Bestehorn-Willmann M, Dobler G, Beers M, Wernike K. In vivo characterization of tick-borne encephalitis virus in bank voles (Myodes glareolus). viruses 2019;11:1069.
- Imhoff M, Hagedorn P, Schulze Y, et al. Review: Sentinels of tick-borne encephalitis risk. Ticks Tick Borne Dis. 2015;6:592- 600.
- Jääskeläinen A, Tonteri E, Pieninkeroinen I, Sironen T, Voutilainen L, Kuusi M, Vaheri A, Vapalahti O. Siberian subtype tick-borne encephalitis virus in Ixodes ricinus in a newly emerged focus, Finland. Ticks Tick borne Dis. 2016;7:216-23.
- Smura T, Tonteri E, Jääskeläinen A, vonTroil G, Kuivanen S, Huitu O, Kareinen L, Uusitalo J, Uusitalo R, Hannila-Handelberg T, Voutilainen L, Nikkari S, Sironen T, Sane J, Castén J, Vapalahti O. Recent establishment of tick-borne encephalitisfoci with distinct viral lineages in the Helsinki area, Finland. Emerging Microbes & Infect. 2019;8:675-683.
- Brandenburg PJ, Obiegala A, Schmuck HM, Dobler G, Chitimia-Dobler L, Pfeffer M. Seroprevalence of Tick-Borne Encephalitis (TBE) Virus Antibodies in Wild Rodents from Two Natural TBE Foci in Bavaria, Germany. Pathogens. 2023 Jan 25;12(2):185. doi: 10.3390/pathogens12020185.
- Knap N, Korva M, Dolinšek V, et al. Patterns of tick-borne encephalitis virus infection in rodents in Slovenia. Zoonotic Dis. 2012;12:236-42.
- Achazi K, Ruzek D, Donoso-Mantke, O, et al. Rodents as Sentinels for the Prevalence of Tick-Borne Encephalitis Virus. Vector-Borne Zoonotic Dis. 2011;11: 641-7.
- Pintér R, Madai M, Horváth G, Németh V, Oldal M, Kemenesi G, Dallos B, Bányai K, Jakab F. Molecular detection and phylogenetic analysis of tick-borne encephalitis virus in rodents captured in the transdanubian region of Hungary, Vector Borne Zoonotic Dis. 2014;14:621-4.
- Zöldi V, Papp T, Rigó K, Farkas J, Egyed L. A 4-Year Study of a Natural Tick-Borne Encephalitis Virus Focus in Hungary. 2010–2013, EcoHealth. 2015;12:174–82.
- Makenov M, Karan L, Shashina N, Akhmentshina M, Zhurenkova O, Kholodilov I, Karganova G, Smirnova N, Grigoreva Y, Yankovskaya Y, Fyodorova M. First detection of tick-borne encephalitis virus in Ixodes ricinus ticks and their rodent hosts in Moscow, Russia. Ticks Tick-Borne Dis. 2019;10:101265.
- Tonteri E, Kipar A, Voutilainen L, et al. The three subtypes of tick-borne encephalitis virus induce encephalitis in a natural host, the bank vole (Myodes glareolus). PLoS ONE. 2013;8:e81214.
- Tonteri E, Jääskeläinen AE, Tikkakoski T, et al. Tick-borne encephalitis virus in wild rodents in winter, Finland (2008- 2009). Emerg Infect Dis. 2011;17:72-5.
- Grzybek M, Tolkacz K, Alsarraf M, Dwużnik D, Szczepaniak K, Tomczuk K, Biernat B, Behnke JM, Bajer A. Seroprevalence of tick-borne encephalitis virus in three species of voles (Microtus spp.) in Poland. J Wildlife Dis. 2020;56;in press.
- Süss J, Gelpi E, Klaus C, et al. Tick-borne encephalitis in naturally exposed monkey (Macaca sylvanus). Emerg Infect Dis. 2007;13:905-7.
- Süss J, Dobler G, Zöller G, et al. Genetic characterization of a tick-borne encephalitis virus isolated from the brain of a naturally exposed monkey (Macaca sylvanus). Int J Med Microbiol. 2008;298(Suppl. 44):295-300.
- Klaus C, Hoffmann B, Beer M, et al. Seroprevalence of tick-borne encephalitis (TBE) in naturally exposed monkeys (Macaca sylvanus) and sheep and prevalence of TBE virus in ticks in a TBE endemic area in Germany. Ticks Tick Borne Dis. 2010;1:141-4.
- Klaus C, Gethmann J, Hoffmann B, et al. Tick infestation in birds and prevalence of pathogens in ticks collected from different places in Germany. Parasitol Res. 2016;115:2729-40.
- Waldenström J, Lundkvist A, Falk KI, et al. Migrating birds and tick-borne encephalitis virus. Emerg Infect Dis. 2007;13:1215- 8.
- Geller J, Nazarova L, Katargina O, et al. Tick-borne pathogens in ticks feeding on migratory passerines in western part of Estonia. Vector Borne Zoonotic Dis. 2013;13:443-8.
- Lommano E, Dvořák C, Vallotton L, Jenni L, Gern L. Tick-borne pathogens in ticks collected from breeding and migratory birds in Switzerland. Ticks Tick Borne Dis. 2014;5:871-82.
- Kazarina A, Japina K, Keiss O, et al. Detection of tick-borne encephalitis virus in I. ricinus ticks collected from autumn migratory birds in Latvia. Ticks Tick Borne Dis. 2015;6:178-80.
- Csank T, Bhide K, Bencúrová E, Dolinská S, Drzewnioková P, Major P, Korytár Ľ, Bocková E, Bhide M, Pistl J. Detection of West Nile virus and tick-borne encephalitis virus in birds in Slovakia, using a universal primer set. Arch Virol. 2016; 161:1679–1683.
- Hubálek Z, Rudolf I. Tick-borne viruses in Europe. Parasitol Res. 2012;111:9-36.
- Mikryukova TP, Moskvitina NS, Kononova YV, et al. Surveillance of tick-borne encephalitis virus in wild birds and ticks in Tomsk city and its suburbs (Western Siberia). Ticks Tick Borne Dis. 2014;5:145-51.
- ECDC (European Centre for Disease Prevention and Control) Epidemiological situation of tick – borne encephalitis in the European Union and European Free Trade Association countries. Stockholm: ECDC; http://www.ecdc.europa.eu/en/ publications/Publications/TBE-in-EU-EFTA.pdf. 2012.
- García-Bocanegra I, Jurado-Tarifa E, Cano-Terriza D, Martínez R, Pérez-Marín JE, Lecollinet S. Exposure to West Nile virus and tick-borne encephalitis virus in dogs in Spain. Transbound Emerg Dis. 2018;65:765-772.
- Salat J, Hunady M, Svoboda P, Strelova L,Strakova P, Fortova A, Paulus M, Ruzek D. Efficacy and immunogenicity of a veterinary vaccine candidate against tick-borne encephalitis in dogs. Vaccine 2023; 41:6150-55.